
Núcleos da base

Os núcleos da base ou núcleos de Giornando, são constituídos por um conjunto de núcleos no cérebro com diferentes estruturas e atividades que atuam como uma unidade funcional. Emitem e recebem projeções entre si e com o córtex cerebral, tálamo e tronco cerebral, e são responsáveis por diversas funções como: coordenação motora, comportamentos de rotina (bruxismo), emoções e cognição.
Os principais constituintes dos gânglios da base são: corpo estriado (núcleo caudado e putâmen), globo pálido, núcleo subtalâmico e substância negra.
Subdivisões anatômicas[editar | editar código-fonte]

ROSTRAL: corpo estriado, globo pálido (GPe and GPi)
CAUDAL: núcleo subtalâmico (STN), substância nigra (SN)
Os núcleos da base são constituídos por diferentes porções de matéria cinzenta, localizados no telencéfalo mais concretamente nas regiões inferiores do cérebro, junto ao tálamo. Ocupam uma posição lateral em ambos os hemisférios cerebrais.
Em termos anatómicos, os núcleos da base podem ser divididos em quatro estruturas, sendo elas: o estriado e globo pálido, as de maiores proporções; e a substância negra e o núcleo subtalâmico, as de menores proporções.[1]
Corpo estriado[editar | editar código-fonte]
O estriado é a maior estrutura dos gânglios da base, e o seu nome deriva da existência de estrias internas. Esta aparência deriva da existência de duas massas de matéria cinzenta separadas por uma região de matéria branca, sendo chamadas respectivamente de núcleo caudado e putâmen.
Esta estrutura é composta principalmente por neurónios GABAérgicos (inibem os neurónios a que se ligam) que projetam dendritos para o globo pálido externo (via indireta) e para o globo pálido interno (via direta). Recebem ainda projeções de neurônios excitatórios do córtex, tálamo e substância negra pars compacta que são essenciais para o movimento.[2]
Globo pálido[editar | editar código-fonte]
O globo pálido é uma estrutura localizada na parte mais interior do estriado. Apesar de parecer uma região homogénea pode ser dividida em duas estruturas funcionais distintas: um segmento interno e outro externo, conhecidos por GPi e GPe respectivamente. Ambas as regiões são constituídas por neurónios GABAérgicos que estão ativos d forma tónica e são responsáveis pela inibição dos movimentos. No entanto, os dois segmentos apresentam funções diferentes uma vez que participam em diferentes circuitos neuronais. O GPe é responsável pela via indireta uma vez que exerce a sua ação emitindo projeções para o núcleo subtalâmico (STN). Por outro lado o GPi recebe ligações não só do estriado mas também do STN, participando na via direta e indireta, e por fim projetando neurónios inibitórios para o tálamo.
Substância negra[editar | editar código-fonte]
A substância negra (ou substância nigra) é uma região do mesencéfalo constituída sobretudo por neurónios dopaminérgicos. Pode ser ainda subdividida em duas estruturas: a substância negra pars reticulata (SNpr) e substância negra pars compacta (SNpc). Estruturalmente, a SNpr pode receber inputs diretamente do estriado (via direta) ou do núcleo subtalâmico via GPe (via indireta), levando a diferentes ações na coordenação motora. Por outro lado, a SNpc é responsável por emitir projeções de neurônios dopaminérgicos para o estriado, onde vai regular a ação dos neurônios inibitórios do GPe, inibindo movimentos involuntários pela via indireta e facilitando os movimentos voluntários pela via direta. A degeneração da substância negra causa o Mal de Parkinson , que pode se manifestar com tremor de repouso, dificuldade em começar ou parar um movimento, desequilíbrio, entre outros.
Núcleo subtalâmico[editar | editar código-fonte]
O núcleo subtalâmico é a única região dos gânglios basais que tem uma ação excitatória através do glutamato. Possui projeções na via indireta nas regiões SNr e GPi.
Neurotransmissores[editar | editar código-fonte]
Os diferentes tipos de neurônios dos núcleos da base sintetizam diferentes neurotransmissores.
| Estrutura | Neurotransmissor | Descrição | Distúrbios |
| Corpo estriado | GABA | Neurônios médios, as células principais, são inibitórios | Doença de Huntington |
| Substância negra | dopamina | A pars compacta da Substância Negra (SNc) primariamente atinge o striatum com seu neurotransmissor (mostrado como uma conexão magenta na conectividade clássica no diagrama abaixo). | Problemas na biossíntese ou transmissão de dopamina podem levar a sérios déficits motores e cognitivos, como ocorre na doença de Parkinson. |
| Globo pálido | GABA | O globo pálido contém um segmento interno e um segmento externo. O segmento interno projeta para o tálamo, ao passo que o segmento externo projeta para o núcleo subtalâmico. | Síndrome de Tourette[3] |
| Núcleo subtalâmico | glutamato | Os neurônios do núcleo subtalâmico excitam neurônios do globo pálido interno. | Lesões no núcleo subtalâmico podem resultar em hemibalismo. |
Seleção de ações: importância dos núcleos basais[editar | editar código-fonte]
Os nucléos da base estão envolvidos em vários circuitos neuronais implicados em funções cognitivas e associativas, motivação, funções motoras e emoções. O estriado é responsável por receber inputs de todas as áreas corticais através do tálamo e projetam principalmente para as áreas pré-frontais e motoras. Estes circuitos revelam subdivisões funcionais (oculomotor, pré-frontal, e giro cingulado) que desempenham funções importantes a nível da atenção e aprendizagem.[4][5]
A ligação estabelecida entre os gânglios da base, tálamo e zonas corticais é responsável por recolher informação das zonas sensoriais por todo o corpo e integrá-las em sinais que posteriormente controlam e corrigem a ação muscular. O estriado é o principal responsável pela inibição das ações desencadeadas pelo córtex e estruturas associadas através das projeções GABAérgicas. Desta forma, os gânglios basais, através do estriado, desempenham um papel importante no controlo de movimentos involuntários e estereotipados. A desregulação desta estrutura está associada a algumas doenças degenerativas como: doença de Parkinson, doença de Wilson, paralisia supranuclear progressiva, doença de Huntington. Estas doenças diferem no tipo de lesão e na sua localização, tendo como sintomas comuns a abulia (incapacidade de iniciar uma ação espontânea) e distonia.[6]
Circuito CGTC[editar | editar código-fonte]
Os gânglios da base são uma estrutura que possui múltiplas ligações diretas com o tálamo e o córtex cerebral, formando um circuito – córtex – gânglios basais – tálamo – córtex (CGTC), que influencia o comportamento motor e também cognitivo.[7][8]
Do córtex, os gânglios da base recebem um input de informação principalmente no corpo estriado. Este estímulo é efetuado através de neurónios glutamatérgicos, excitatórios, podendo atuar em ambos recetores dopaminérgicos D1 e D2 existentes no estriado. Daí, os neurónios projetam-se para o globo pálido interno/substância negra pars reticulata (GPi/SNr) ou pálido externo (GPe), formando a 'via direta 'ou a' indireta'.[9][10]
A atividade do corpo estriado é modulada pela substância negra pars compacta (SNc), cujas projeções dopaminérgicas e a sua maior ou menor libertação fásica de dopamina mantem as vias em equilíbrio.[11][12]
_e_indireta_(NoGO).jpg)
A transmissão da informação pela via direta (GO) é iniciada pela ativação dos recetores D1 no estriado. Daí, existe a projeção sucessiva de neurónios GABAérgicos (inibitórios) para o GPi/SNr, ocorrendo o silenciamento da atividade tónica dos seus neurónios inibitórios; de seguida, há a projeção para o tálamo. Desta forma, o input excitatório proveniente do córtex (glutamatérgico) irá desinibir os neurónios talâmicos, que por sua vez estimulam o córtex. Esta via resulta assim na facilitação da atividade muscular, por estimulação da via corticoespinhal lateral pelo córtex.[7][8]
A via indireta (NoGO) envolve a passagem de informação pelo GPe, seguindo para o GPi/SNr e núcleos subtalâmicos, sendo ativada pela estimulação dos recetores D2 no estriado. O GPe envia um sinal inibitório para o núcleo subtalâmico (STN), que por sua vez possui uma ligação excitatória com o complexo GPi/SNr. Desta forma, a ativação da via indireta leva ao aumento do output pelo GPi, inibindo o tálamo, resultando num movimento menor e mais lento pela diminuição da estimulação do córtex.[9][13]
O STN recebe também projeções glutamatérgicas provenientes do córtex frontal e motor, que irão aumentar também a atividade do GPi através de uma excitação geral e rápida. Esta via é denominada hiperdireta e permite a prevenção de uma resposta prematura, sendo particularmente importante em situações de extenso conflito entre ativação das diferentes vias através da inibição de atividade motora indesejada.[14]
Assim, como resultado da coordenação entre excitação, inibição e desinibição, perante a ativação da via direta (GO) obtemos uma ação motivada, através da estimulação do córtex (circuito de feedback positivo). Porém, quando a via indireta (NoGO) se encontra mais ativa, existe supressão da atividade do córtex (circuito de feedback negativo). Desta forma, é possível a escolha entre possíveis ações a realizar, podendo facilitar uma ação adaptativa e ao mesmo tempo suprimir as ações competitivas, mantendo o equilíbrio entre a estimulação e inibição motora.[7][9][12]
Disfunção dos núcleos da base[editar | editar código-fonte]
Doença de Parkinson[editar | editar código-fonte]
A Doença de Parkinson é uma doença idiopática caracterizada pela destruição dos neurônios dopaminérgicos da substância negra e que tem como consequência a redução drástica dos níveis de dopamina no corpo estriado dos pacientes; os neurônios dopaminérgicos enervam as principais estruturas dos gânglios da base (corpo estriado, os segmentos externo e interno do globo pálido, o núcleo subtalâmico, substância negra), assim como os alvos das suas projeções (tálamo e córtex motor, por exemplo) servindo como moduladores dos circuitos corticoestriatais.[15][16][17]
A perda de dopamina nestas zonas compromete o delicado equilíbrio entre inibição tónica e “desinibição” dos circuitos motores e da seleção consciente de um determinado movimento. Níveis saudáveis de dopamina potenciam a via direta (GO) e impedem a via indireta (NoGO), uma vez que os neurónios da via direta expressam recetores de dopamina D1 (ativadores) e os neurónios da via indireta expressam recetores de dopamina D2 (desactivadores). Por outro lado, baixos níveis de dopamina revertem esse efeito: impedem a via GO e favorecem a via NoGO, resultando numa excessiva inibição tálamo-cortical.[16][17]

Esta disfunção do circuito que os gânglios da base mantêm entre o córtex e o tálamo motor é a base dos sintomas motores clássicos parkinsonianos: tremores, hipocinesia (lentidão e dificuldade em iniciar movimentos), rigidez muscular e instabilidade postural.[15]
Tal como o circuito motor, outros importantes circuitos que envolvem os gânglios da base, e que estão portanto dependentes da dopamina, são afetados na Doença de Parkinson; são estes os circuitos: oculomotor, associativo, límbico e orbito frontal, responsáveis por variadas funções como a atenção, aprendizagem, memória e emoções. Curiosamente, todas estas funções são afetadas na Doença de Parkinson, manifestando-se como sintomas não-motores de disfunções da fala, funções autônomas, problemas a nível da memória e raciocínio, podendo mesmo chegar a demência em alguns casos. Apesar de, cientificamente, a relação entre o circuito motor dos núcleos da base e a Doença de Parkinson ser a mais estudada e reconhecida, a relação entre os circuitos que envolvem os núcleos da base e os sintomas não-motores da doença não é ainda completamente compreendida. No entanto, novos estudos começam a apontar para esta relação como causa provável dos sintomas não-motores desta doença.[17][18][19]
O tratamento clássico da Doença de Parkinson é a administração de L-DOPA (ou levodopa, um agonista da dopamina) para restaurar os níveis deste neurotransmissor. Este tratamento é capaz de reverter os efeitos da morte dos neurónios dopaminérgicos, ainda que doses elevadas possam causar uma ativação exagerada da via GO distorcendo a noção de recompensa do indivíduo, levando a novos sintomas de obsessão e comportamentos compulsivos, problemas com o jogo entre outros.

Síndrome de Tourette[editar | editar código-fonte]
A Síndrome de Tourette é uma doença neuropsiquiátrica idiopática caracterizada pela ocorrência crónica de variados tiques (ações involuntárias e repetitivas) motores (piscar os olhos, torcer o nariz, abrir a boca etc.) e/ou vocais (grunhidos, guinchos etc.). Outros sintomas como comportamentos obsessivos, falta de atenção e impulsividade estão frequentemente associados à Síndroma de Tourette. Como já foi mencionado acima, a causa desta doença é desconhecida mas estes sintomas, relacionados com a incapacidade de controlar movimentos musculares, noção distorcida de recompensa, repetição e rapidez nos movimentos, parecem apontar para uma disfunção a nível dos núcleos da base e do circuito motor e/ou frontocortical. De facto, a hipercinesia característica desta doença opõe-se à hipocinesia característica da Doença de Parkinson pelo que, ao contrário do que se passa nesta última, a Síndrome de Tourette parece ser devido a excessiva ativação do corpo estriado. Este fenómeno pode ser devido à libertação de maiores quantidades de dopamina, maior sensibilidade por parte dos recetores de dopamina D1 ou maior enervação dopaminérgica. Como consequência, há uma exagerada ativação da via GO em relação à via NoGO facilitando a execução de certos movimentos motores e impedindo a inibição de movimentos concorrentes ao movimento desejado dando origem aos tiques característicos.[17][20][21]
Os tiques podem ter início aquando da ativação inapropriada e repetida de um conjunto de nervos do corpo estriado que, devido a um ciclo vicioso de ativações repetidas, origina um mecanismo de plasticidade que facilita esse mesmo movimento, criando um hábito e consolidando o tique.[20][21]
O tratamento da Síndrome de Tourette dá-se à base de bloqueadores dos recetores de dopamina D2, que irá promover a via NoGO em detrimento da via GO e assim evitar a “desinibição” de movimentos involuntários. O bloqueio destes recetores causa também mecanismo de plasticidade nos neurónios da via NoGO que aumenta a capacidade de aprendizagem negativa (ou “aprendizagem NoGO”) possibilitando uma abordagem extra no tratamento permanente dos tiques, onde a ausência destes seria recompensada de modo a “desabituar” o paciente dos tiques.[17]
Imagens adicionais[editar | editar código-fonte]
-
 Secção horizontal através do telencéfalo
Secção horizontal através do telencéfalo -
 Cortes coronais dos núcleos da base
Cortes coronais dos núcleos da base -
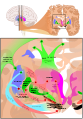 Vias inibitórias e excitatórias
Vias inibitórias e excitatórias


Referências
- ↑ Ullsperger, M., Danielmeier, C. & Jocham, G. Neurophysiology of performance monitoring and adaptive behavior. Physiol Rev 1, 35–79 (2014).
- ↑ Fix, J. D. in Neuroanatomy 274–281 (Lippincott Wiliams & Wilkins, 2008)
- ↑ «Síndrome de Tourette e globo pálido». Rede Record. Consultado em 27 de janeiro de 2009. Arquivado do original em 28 de março de 2009
- ↑ John, R. et al. Conditional Routing of Information to the Cortex: A Model of the Basal Ganglia’s Role in Cognitive Coordination. Psychol. Rev. 117, 541–74 (2010)
- ↑ Chakravarthy, V. S., Joseph, D. & Bapi, R. S. What do the basal ganglia do? A modeling perspective. Biol. Cybern. 103, 237–53 (2010).
- ↑ Hollerman, J., Tremblay, L. & Schultz, W. Involvement of basal ganglia and orbitofrontal cortex in goal-directed behaviour. Prog Brain Res 126, 193–215 (2000).
- ↑ a b c Goldberg, J. H., Farries, M. A. & Fee, M. S. Basal ganglia output to the thalamus: still a paradox. Trends Neurosci. 36, 695–705 (2013)
- ↑ a b The MIT Encyclopedia of the Cognitive Sciences (MITECS). (A Bradford Book, 2001)
- ↑ a b c Schroll, H. & Hamker, F. H. Computational models of basal-ganglia pathway functions: focus on functional neuroanatomy. Front. Syst. Neurosci. 7, 122 (2013)
- ↑ Wichmann, T. & DeLong, M. R. Functional and pathophysiological models of the basal ganglia. Curr. Opin. Neurobiol. 6, 751–758 (1996)
- ↑ Mink, J. W. The basal ganglia: focused selection and inhibition of competing motor programs. Prog. Neurobiol. 50, 381–425 (1996)
- ↑ a b Yin, H. H. & Knowlton, B. J. The role of basal ganglia in habit formation. Nat. Rev. Neurosci. 7, 464–476 (2006)
- ↑ Onla-or, S. & Winstein, C. J. Function of the “direct” and “indirect” pathways of the basal ganglia motor loop: evidence from reciprocal aiming movements in Parkinson’s disease. Cogn. Brain Res. 10, 329–332 (2001)
- ↑ Nambu, A., Tokuno, H. & Takada, M. Functional significance of the cortico-subthalamo-pallidal “hyperdirect” pathway. Neurosci. Res. 43, 111–7 (2002)
- ↑ a b Jankovic, J. Parkinson’s disease: clinical features and diagnosis. J. Neurol. Neurosurg. Psychiatry 79, 368–76 (2008)
- ↑ a b Obeso, J. a et al. Functional organization of the basal ganglia: therapeutic implications for Parkinson’s disease. Mov. Disord. 23 Suppl 3, S548–59 (2008)
- ↑ a b c d e Maia, T. V, & Frank, M. J. (2011). From reinforcement learning models to psychiatric and neurological disorders. Nature neuroscience, 14(2), 154–62.
- ↑ Sawamoto, N. et al. Cognitive slowing in Parkinson disease is accompanied by hypofunctioning of the striatum. Neurology 68, 1062–8 (2007)
- ↑ Bodden, M. E., Dodel, R. & Kalbe, E. Theory of mind in Parkinson’s disease and related basal ganglia disorders: a systematic review. Mov. Disord. 25, 13–27 (2010)
- ↑ a b Mink, J. W. Neurobiology of Basal Ganglia and Tourette Syndrome: Basal Ganglia circuits and thalamocortical outputs. Adv. Neurobiol. 99, 89–98 (2006)
- ↑ a b Mink, J. W. Basal Ganglia Dysfunction in Tourette’ s Syndrome : A New Hypothesis. 25, 190–198 (2001)