
Lophiiformes
| Lophiiformes peixes-pescadores, tamboris | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ocorrência: 130–0 Ma
Cretáceo inferior – recente | |||||||||||||||
| Classificação científica | |||||||||||||||
| |||||||||||||||
| Subordens | |||||||||||||||


(B) Cryptopsaras couesii, 34.5 mm SL
(C) Himantolophus appelii, 124 mm SL
(D) Diceratias trilobus, 86 mm SL
(E) Bufoceratias wedli, 96 mm SL
(F) Bufoceratias shaoi, 101 mm SL
(G) Melanocetus eustalus, 93 mm SL
(H) Lasiognathus amphirhamphus, 157 mm SL
(I) Thaumatichthys binghami, 83 mm SL
(J) Chaenophryne quasiramifera, 157 mm SL.

Lophiiformes (Lofiiformes)[1] é uma ordem de peixes ósseos que agrega um conjunto muito diverso de peixes marinhos, com cerca de 322 espécies repartidas por 5 subordens e 16-18 famílias,[2] caracterizados na maioria das espécies pela presença na parte anterior da cabeça de um aparato pra atração de presas (o illicium com a sua esca, formando um isco, daí serem conhecidos por peixes-pescadores) , estrutura resultante da modificação do primeiro raio da barbatana dorsal.[3][4] Outra característica diferenciadora é a presença de dimorfismo sexual extremo, especialmente na subordem Ceratioidei.[5] A ordem engloba maioritariamente espécies da zona afótica das águas profundas de todos os oceanos,[6] mas também estão presentes espécies pelágicas e bentónicas dos recifes de coral e das águas costeiras. Apenas se conhece uma espécie desta ordem (Antennarius biocellatus) que vive em águas salobras ou doces. Algumas das espécies são bem conhecidas pela sua morfologia exótica, nomeadamente os peixes-morcego (Ogcocephaloidei), os peixes-sapo (Antennarioidei) e os peixes-diabo (Ceratioidei).[3] Algumas espécies da família Lophiidae, que inclui o tamboril, apresentam interesse comercial.
Descrição
[editar | editar código-fonte]A ordem Lophiiformes agrupa os peixes ósseos (Osteichthyes) teleósteos[7] conhecidos pelo nome comum de peixes-pescadores ou, mais restritamente, tamboris. São predadores cujo nome comum resulta do seu modo incomum de predação, no qual a utilização de um isco formada pelo alongamento de um raio modificado da barbatana dorsal (o ilício) que termina numa estrutura bolboas (a esca ) que serve para atrair outros peixes, a qual pode ser luminescente.
Quando presente, a bioluminescência que ocorre em muitas espécies é produzida por bactérias simbióticas, que se crê sejam adquiridas da águas do mar através da criação na esca de condições ambientais e tróficas que favorecem a sua fixação e crescimento,[8][9] formando um microbioma específico dentro e em torno da cavidade da esca.
Alguns Lophiiformes são também notáveis por apresentarem dimorfismo sexual muito acentuado, ou mesmo extremo, que, nalguns casos atinge a simbiose sexual, com o macho muito mais pequeno do que a fêmea. Esta característica é mais marcada na subordem Ceratiidae, um grupo de peixes da região abissal, na qual algumas espécies têm machos que são várias ordens de magnitude menores do que as correspondentes fêmeas.[10]
O grupo tem distribuição natural cosmopolita, ocorrendo em todos os oceanos. Entre os membros desta ordem contam-se espécies pelágicas, que ocorrem longe dos fundos marinhos, e espécies bentónicas cuja distribuição está condicionada à proximidade imediata dos fundos. Algumas vivem no mar profundo (como os membros da família Ceratiidae), enquanto outras ocorrem na plataforma continental (como os peixes-rã da família Antennariidae e os tamboris e os peixes-ganso da família Lophiidae).
As formas pelágicas são mais frequentemente lateralmente comprimidas, enquanto as formas bentónicas são frequentemente extremamente comprimidas dorso-ventralmente, em geral apresentando grandes bocas apontadas para cima.
Morfologia
[editar | editar código-fonte]

Todos os membros da ordem Lophiiformes são carnívoros, apresentando consequentemente uma anatomia que resulta da adaptação para a captura de presas, o que se traduz na presença de coloração mimética e de bocas e dentes adaptados à captura e ingestão de peixes, a que acresce a presença do ilício .
Variando em coloração do cinza escuro ao castanho escuro, as espécies de águas profundas apresentam grandes cabeças terminadas em enormes bocas em forma de meia-lua, tendo na sua frente numerosos dentes longos e afiados em forma de garras, angulados para dentro, permitindo uma captura eficiente das presas. O comprimento corporal destes peixes varia geralmente de 2 a 18 cm, mas com algumas espécies a atingir os 100 cm,[11] mas a variação é devida em grande parte ao muito acentuado dimorfismo sexual, com as fêmeas muito maiores do que os machos.[12]
Os peixes-rã e outras espécies de águas pouco profundas são predadores que caçam por emboscada e muitas vezes aparecem camuflados como rochas, esponjas ou algas marinhas, apresentando coloração variada em função das condições do habitat, com frequentes casos de cores vivas e pele com protuberâncias e excrescências multicolores de morfologia complexa.[13]
Apesar dessa variabilidade de formas e de habitats, a ordem Lophiiformes é caracterizada por exibir variações morfológicas típicas e únicas, principalmente na boca e na estrutura da barbatana dorsal, que é espinhosa e sofre a adaptação mais distintiva deste agrupamento taxonómico.
Para além das adaptações específicas atrás apontadas, estes peixes caracterizam-se pela presença de aberturas branqueais restritas a aberturas tubulares, pequenas e alongadas, situadas próximas da barbatana peitoral, e por apresentarem os primeiro e segundo centros urais fundidos com o primeiro centro pleural, formando uma placa que surge de um único meio-centro. Os ossos cranianos epióticos são separados dos parietais e localizam-se na linha média posterior à região supra-occipital. Os raios radiais das barbatanas peitorais são alongados e estreitos, onde o raio mais ventral é consideravelmente expandido distalmente.
Além disso, uma das sinapomorfias do grupo é a produção de ovos com dupla bainha, envoltos por uma massa gelatinosa que permite que flutuem agrupados.[6][14] A estrutura que envolve os ovos é uma fina película de material gelatinoso e transparente, com 50 a 75 cm de largura e de 6 a 7,5 m de comprimento. Os ovos estão envoltos por uma bainha individual e cada um está inserido na sua própria cavidade. As massas de ovos são libertadas no mar, flutuando livremente. As larvas nadam livremente e apresentam a pélvis alargada em filamentos.
Para além disso, estes peixes apresentam fusão da primeira vértebra com o crânio. As barbatanas pélvicas, quando presentes, estão posicionadas à frente das peitorais, com um espinho e de quatro a cinco raios moles. Como adaptação à ingestão de grandes presas, há ausência de costelas, permitindo a distensão do estômago para acomodar grandes volumes.[6]
- Boca e dentes
A maioria das espécies apresenta uma boca extremamente alargada que se estende por toda a circunferência anterior da cabeça, provida de faixas de dentes, inclinados para dentro, alinhados sobre ambas as mandíbulas. Os dentes podem ser rebaixados de modo a não oferecer impedimento para um objeto deslizar em direção ao estômago, mas evitar que a presa, depois de entrar, possa escapar da boca.[15]
Estes peixes são capazes de distender a mandíbula e o estômago, uma vez que seus ossos são finos e flexíveis, chegando a tamanhos enormes, o que lhe permite engolir presas até duas vezes maiores que o seu corpo inteiro.[16]
- Illicium (ou ilício) e esca
A barbatana dorsal dos membros desta ordem é formada por seis lepidotríquias em forma de espinhos, sendo que as três frontais são cefálicas e modificadas para formar o illicium (ou ilício), um apêndice fino e alongado que termina numa estrutura bolbosa designada por esca (ou isca).
O ilício constitui uma estrutura de atração da presa para a boca, semelhante a uma cana de pesca, com o isco localizado frente à enorme boca do peixe. Esta estrutura, exclusiva deste grupo, levou ao nome comum de peixes-pescadores.
O termo «ilício» deriva do latim illicĭu-, «atrativo», a mesma origem etimológica do termo português ««ilício» utilizado para designar o acto ou crime de iliçar, ou seja de dolosamante enganar.
A estrutura constitui assim um ardil, ou isco, destinado a atrair as presas para uma posição fontal à boca, levando a que sejam facilmente engolidas por uma aspiração rápida da água acompanhada por uma muito rápida projecção do peixe para diante com a enorme boca distendida. Em alguns casos esta estrutura está ausente dos machos, especialmente em espécies que exibem simbiose sexual, com os machos essencialmente parasitando o metabolismo das fêmeas sobre as quais se fixam. A estrutura é constituída por duas partes distintas:
- O illicium, ou ilício propriamente dito, de forma linear, podendo nalguns casos ser muito longo e flexível, quase como uma linha, embora noutros casos seja curto e engrossado. A estrutura é formada a partir dos três raios frontais da barbatana dorsal, sendo que um deles é modificada por alongamento, emergindo como um verdadeiro pedúnculo na região cefálica (a cabeça). No extremo distal deste pedúnculo localiza-se o crescimento carnudo designado por esca (ou isca);
- A esca, ou isca, é uma estrutura bulbosa, carnuda, com morfologia complexa e muito variável entre géneros e espécies, que pode ter diversos tipos de apêndices posteriores ou posterolaterais. Nalguns casos apresenta na sua superfície dentículos ósseos. A esca das espécies de grande profundidade apresenta em geral bioluminescência, resultado da presença de bactérias luminescentes sobre a sua superfície. Para permitir a fixação e a formação de colónias bacterianas, a esca apresenta uma «glândula luminosa» constituída por uma concavidade aberta por um poro à água do mar, onde as bactérias luminescentes se agrupam.
Na maioria das espécies ceratióides (subordem Ceratioidei) as fêmeas adultas apresentam escas luminescentes na ponta de um longo ilício. Tem-se especulado que este órgão para além de servir o propósito óbvio de atrair presas nos ambientes escuros do mar profundo, também serve para chamar a atenção dos machos para as fêmeas, facilitando o acasalamento.
Nessas espécies, a fonte de luminescência são bactérias simbióticas que habitam dentro e ao redor da esca, envoltas num refletor em forma de copo contendo cristais, provavelmente de guanina. Apenas um pequeno conjunto de espécies de bactérias simbiontes luminescentes se podem associar a estes peixes de águas profundas.[3]
Em algumas espécies, as bactérias recrutadas para a esca são incapazes de luminescência quando separadas do peixe, sugerindo que desenvolveram um relação simbiótica, sendo as bactérias incapazes de sintetizar autonomamente todos os compostos necessários para a luminescência. Estas espécies dependem assim do metabolismo dos peixes para completar o processo. A microscopia eletrónica das bactérias presentes em algumas espécies de peixes revela que são bactérias Gram-negativas em forma de bastonete (bacilos) sem cápsulas, esporos ou flagelos. Apresentam paredes celulares de dupla camada e mesossomas.
Um poro liga a esca com a água do mar, o que facilita a remoção das bactérias mortas e dos resíduos celulares, e permite que o pH e a tonicidade do meio de cultura permaneçam constantes. Essas condições, a que acresce a temperatura constante da zona batipelágica habitada por estes peixes, é crucial para a viabilidade a longo prazo das culturas bacterianas presentes na esca.[17][18]
Como a «glândula luminosa» está sempre aberta para o exterior através de um poro, é possível que o peixe adquira as bactérias a partir da água do mar. No entanto, parece que cada espécie de peixe está associada à uma espécie particular de bactéria, e essas bactérias nunca foram encontradas isoladas na água do mar. Para explicar estas observações, foi teorizado que a esca descarrega bactérias durante a desova e as bactérias são assim transferidas para os ovos.[18][19]
Algumas evidências mostram que alguns Lophiiformes adquiriram os seus simbiontes bioluminescentes do ambiente local. Materiais genéticos das bactérias simbiontes são encontrados perto dos peixes, indicando que o peixe e as suas bactérias associadas provavelmente não evoluíram juntos e as bactérias fazem jornadas difíceis para entrar no hospedeiro.[3] Num estudo conduzido sobre uma espécie da subordem Ceratioidei no Golfo do México, os investigadores notaram que os micróbios bioluminescentes que foi possível estabelecer com certeza como associados ao hospedeiro não estão presentes nos espécimes larvais e durante o desenvolvimento inicial do hospedeiro. Tal leva a concluir que os ceratióides provavelmente adquirem os seus simbiontes bioluminescentes a partir da água do mar.[20]
A espécie Photobacterium phosphoreum, e os restantes membros do clado que inclui Photobacterium kishitanii, constituem o principal, ou único, grupo de simbiontes bioluminescentes de várias famílias de peixes luminosos do mar profundo.[21]
Sabe-se que a composição genética das bactérias simbiontes sofreu alterações desde que estas se associaram ao seu hospedeiro.[3] Em comparação com os seus parentes de vida livre, os genomas destes simbiontes são reduzidos em mais de 50% do seu tamanho. São encontradas reduções nas vias de síntese de aminoácidos e na capacidade de utilizar diversos açúcares. No entanto, os genes envolvidos na quimiotaxia e na motilidade, que são considerados úteis apenas fora do hospedeiro, são retidos no genoma. O genoma do simbionte contém um número muito alto de pseudogenes e mostra expansões massivas de elementos transponíveis. O processo de redução do genoma ainda está em andamento nessas linhagens simbiontes, e a progressiva redução do genoma leva a uma crescente dependência em relação ao hospedeiro.[22]
Filogenia e sistemática
[editar | editar código-fonte]





Filogenia
[editar | editar código-fonte]Um estudo filogenético do genoma mitocondrial de um conjunto de Lophiiformes sugeriu que este grupo diversificou-se num curto período de tempo, compreendido entre o início e meados do Cretáceo, entre 130 e 100 milhões de anos atrás.[23]
Os primeiros registos fósseis de Lophiiformes são provenientes de depósitos datados do Eoceno, onde surgiram géneros como Lophius e Antennarius.[24]
Os Lophiiformes foram classificados ao longo dos anos como parte de diferentes agrupamentos taxonómicos, sendo durante muito tempo atribuídos aos Paracanthopterygii[6][25] juntamente com outras ordens de peixes sem espinhos nas barbatanas, tais como os pertencentes às antigas ou actuais ordens Scorpaeniformes, Moroniformes, Acanthuriformes e Spariformes. Naquela classificação, os peixes-rã (Batrachoidiformes) eram considerados como os parentes filogeneticamente mais próximos deste grupo. Contudo, a investigação filogenética demonstrou que os Paracanthopterygii daquela época eram um grupo parafilético, o que levou ao seu desmembramento.
Os actuais Lophiiformes também já foram alocados como subordem dos Perciformes, mas estudos de genética molecular recentes mostram claramente que estes peixes devem ser agrupados com os parentes da perca (Percomorphaceae) e que estão intimamente relacionados com os parentes do peixe-balão (Tetraodontiformes) e do peixe-pau (Caproidae).[26][27]
Os Lophiiformes são atualmente considerados como uma ordem pertencente à superordem Percomorpha. Esta, contudo, permanece uma superordem controversa dentro do agrupamento taxonómico dos Vertebrata.[6]
Sistemática
[editar | editar código-fonte]As mais recentes classificações subdividem a ordem Lophiiformes em cinco subordens (Lophioidei, Antennarioidei, Chaunacoidei, Ogcocephaloidei e Ceratioidei), que compartilham modificações evolutivas distintas.[28]
A subordem Lophioidei é considerada o grupo irmão mais basal das outras subordens. As subordens Antennarioidei, Chaunacoidei, Ogocephaloidei e Ceratioide compartilham a redução do tamanho do ovo e do estágio larval, redução do número dos espinhos dorsais de três para menos. As subordens Chaunacoidei, Ogocephaloidei e Ceratioidei compartilham um espinho dorsal secundário localizado abaixo do tegumento da cabeça e a ausência de filamentos branquiais no primeiro arco branquial. As subordens Ogocephaloidei e Ceratioidei compartilham uma redução do segundo espinho dorsal e a ausência do terceiro espinho dorsal.[6]
Theodore Pietsch e Christopher Kenaley, numa recente revisão deste grupo, reconhecem 18 famílias, 65 géneros e 322 espécies. A subordem Lophioidei de peixes das águas profundas, que inclui o tamboril, agrupa mais de metade de todas as espécies de Lophiiformes conhecidas.[29]
Na sua presente circunscrição taxonómica, as subordens e respectivas famílias são as seguintes:[6]
- Subordem Lophioidei
- Subordem Antennarioidei
- Subordem Chaunacoidei
- Subordem Ogcocephaloidei
- Subordem Ceratioidei
As relações filogenéticas entre as diferentes subordens são as constantes do seguinte cladograma:[30]
| Lophiiformes |
| ||||||||||||||||||||||||
Subordem Lophioidei
[editar | editar código-fonte]
Esta subordem apresenta diversas características distintivas, entre as quais a presença de barbatanas pélvicas modificadas e de uma barbatana dorsal espinhosa, localizada atrás da cabeça e com um a três espinhos. O corpo é desprovido de escamas.[6]
A cabeça é grande, achatada e larga, com dentes bem desenvolvidos. Estes peixes apresentam uma série de dentículos ou excrescências cutâneas "franjadas" ao redor da mandíbula e dispersa pelo resto do corpo, principalmente sobre a cabeça.
O ilício apresenta uma esca carnosa na sua extremidade, que atua como uma isco, atraindo presas ao alcance de sua grande boca.[6]
Esta subordem é monotípica, tendo como única família a família Lophiidae, a qual agrupa 4 géneros de peixes das regiões profundas dos oceanos Atlântico, Índico, Pacífico e Ártico, ocorrendo ainda no mar Mediterrâneo.
O grupo inclui o tamboril, um peixe objecto de importante pescaria comercial nas regiões temperadas de todos os oceanos.[31]
Subordem Antennarioidei
[editar | editar código-fonte]
Entre as características compartilhadas pelos integrantes desta subordem, conta-se ter o espinho da barbatana dorsal composto por três espinhos cefálicos, sendo o primeiro o illicium modificado. O terceiro espinho da barbatana dorsal exibe expansões dorsais intensamente comprimidas.
Os membros desta subordem podem ser bentónicos ou pelágicos, e ter os corpo cobertos com pele frouxa e nua ou com dentículos, variando muito entre as famílias.[6]
A subordem Antennarioidei apresenta quatro famílias:
Com uma única excepção, as espécies que integram esta subordem são marinhas, habitando os mares tropicais e subtropicais (com exceção do mar Mediterrâneo). Algumas das famílias podem ser encontradas nas costas da Austrália, Nova Guiné e Ilhas da Indonésia.[6] Pertence a esta subordem a única espécie de Lophiiformes, a espécie Antennarius biocellatus, que se conhece viver em águas salobras ou doces.[32]
Subordem Chaunacoidei
[editar | editar código-fonte]
Esta subordem é monotípica, apresentando uma única família, a família Chaunacidae, que agrupa os peixes conhecidos pelo nome comum de sapos marinhos (sea toads em inglês).[6]
Apresentam corpo globoso, com epiderme coberta por dentículos. Nesta família apenas o illicium se apresenta como raio dorsal espinhoso. A coloração varia entre o rosa e o alaranjado escuro.[6]
As espécies que integram esta subordem são peixes marinhos que habitam os oceanos Atlântico, Pacífico e Índico a profundidades que variam de 90 m a mais de 2000 m.
Subordem Ogcocephaloidei
[editar | editar código-fonte]
Essa subordem é monotípica, integrando apenas a família Ogcocephalidae, cujas espécies são popularmente conhecidos por peixes-morcego (em inglês: "bat-fishes").[6]
Apresentam corpo achatado ventralmente, com illicium diminuto e o remanescente da espinha dorsal secundária presente. Nadam sobre o fundo por intermédio das suas largas barbatanas peitorais, que se assemelham a braços. As barbatanas pélvicas são reduzidas.[6]
São espécies bentónicas de profundidade, habitando em fundos de 1500 a 3000 metros, com pelo menos uma espécie que chega a atingir 4000 m de profundidade.[6]
Ocorrem nas regiões tropicais de todos os oceanos e estão presentes em vários mares subtropicais (com exceção do mar Mediterrâneo).
Subordem Ceratioidei
[editar | editar código-fonte]
Esta subordem é a mais diversa de entre os Lophiiformes. Os seu integrantes caracterizam-se geralmente pela ausência de escamas, mas podem apresentar espinhos e placas cutâneas.
O dimorfismo sexual nesta subordem é tão acentuado que uma fêmea pode atingir um tamanho três a treze vezes superior ao do macho, variando de acordo com a família. Os adultos apresentam uma aparência muito distinta quando comparados às fêmeas. Entre outras características da subordem, pode-se citar a presença de ossos denticulares nos machos anões, com a presença de illicium a ocorrer exclusivamente nas fêmeas.
Os machos destas espécies podem apresentar hábito de vida livre ou parasitário sobre fêmeas, que pode ser obrigatório ou facultativo (alternando entre ambos). O estado parasitário dos machos ocorre em pelo menos quatro das onze famílias, nas quais os machos se fixam em definitivo ao corpo das fêmeas, sofrendo de seguida uma metamorfose para o estágio parasitário, alimentando-se do sangue da fêmea com atrofia de todos os seus órgãos, com excepção das gónadas.[6]
Presente nos oceanos Atlântico, Índico e Pacífico, a subordem é constituída pelas seguintes 11 famílias:
- Centrophrynidae
- Ceratiidae
- Himantolophidae
- Diceratiidae
- Melanocetidae
- Thaumatichthyidae
- Oneirodidae
- Caulophrynidae
- Neoceratiidae
- Gigantactinidae
- Linophrynidae
Ecologia
[editar | editar código-fonte]Modo de vida e comportamento alimentar
[editar | editar código-fonte]Os peixes da ordem Lophiiformes são predadores de emboscada inativos (em inglês: "sit-and-wait predators") que atraem a presa com uma isca formada por adaptação de um raio modificado da barbatana dorsal (o illicium) equipado, geralmente, com uma estrutura bulbosa no seu extremo distal (a esca) frente à boca, que no caso das espécies de profundidade é geralmente um órgão bioluminiscente.[33]
Em geral, os Lophiiformes alimentam-se de várias espécies de peixes, porém há dois tipos principais: os peixes bento-pelágicos, nadando quase continuamente perto do fundo; e os peixes bentónicos, descansando no fundo oceânico sem deslocamento ativo.[34] Como os peixes deste grupo possuem pouca habilidade natatória, passam o maior parte da sua vida aguardando, no caso dos bentónicos enterrados no substrato não consolidado dos fundos ou camuflados sobre o fundo ou entre algas, ou, no caso dos bento-pelágicos, pairando na água, em ambos os casos mexendo a isca para frente e para trás, tentando atrair a potencial presa para a boca, especialmente quando a presa esteja localizada na frente ou acima do seu corpo. Este comportamento alimentar assenta na aproximação da presa ao illicium e por consequência à região frontal do indivíduo.
Um género da família Linophrynidae, o género Linophryne (inglês: "netdevils"), desenvolveu um barbilhão bioluminescente para complementar a sua esca de bioluminescência bacteriana.[34] Esta atração da presa para a esca e outros órgãos bioluminescentes é provavelmente baseada na resposta fototáxica positiva de muitos invertebrados e peixes planctónicos.[35]
A esca não serve apenas para atração de presa, mas também para a comunicação intra-específica, ou seja entre indivíduos de mesma espécie.[36] Esta comunicação é particularmente importante no acasalamento de espécies que habitam a região afótica, onde a densidade populacional das espécies é baixa e os parceiros sexuais difíceis de encontrar.
As espécies da ordem Lophiiformes que ocorrem nas águas profundas vivem num meio com pouco fornecimento alimentar, consequência da diminuição da densidade de partículas orgânicas com o aumento da profundidade.[35] Nessas regiões, as fêmeas alimentam-se de organismos de tamanhos variáveis, desde pequenos crustáceos até lulas e peixes relativamente grandes.[37]
A boca grande, o estômago extensível e um comportamento alimentar de sucҫão (em algumas espécies), permitem a ingestão de presas de relativamente grande tamanho, ao mesmo tempo que aumentam a probabilidade de captura de alimento, fator importante num ambiente de baixa biomassa.[35]
Para defender-se de predaҫão de piscívoros (predadores de peixes) de maiores dimensões, os Lophiiformes emitem uma série de lampejos curtos da esca visando para assustar, ou pleo menos e intimidar, o potencialpredador. Em algumas espécies função semelhante é conseguida com recurso à extrusão de um fluido bioluminescente, que confunde os potenciais predadores .[35]
Reprodução
[editar | editar código-fonte].gif)
A dificuldade em estudar a reprodução de espécies que se abrigam em regiões profundas dificulta a disponibilidade de informações sobre a cópula desses animais. Mesmo em relação a espécies bem conhecidas e com valor económico como pescaria comercial, como por exemplo as pertencentes ao género Lophius (os tamboris), da subordem Lophioidei, pouco se sabe sobre os hábitos reprodutivos. Ainda assim, sabe-se que as fêmeas deste género produzem e liberam os ovos em forma de uma grande massa gelatinosa, agregada e flutuante.
Os lofiídeos destacam-se pela proteção e dispersão dos ovos, que são envolvidos por uma secreção produzida pela fêmea durante a desova que forma uma grande massa gelatinosa flutuante, em formato de tira, que pode exceder dez metros de comprimento e conter mais de um milhão de ovos. Tal massa visa protegê-los de predadores e facilita a emissão de um grande número de ovos, numa clara estratégia reprodutiva do tipo r.
A migração esta presente entre os Lophiiformes, como por exemplo no género Lophius no qual os indivíduos adultos vão para águas profundas durante a desova As tiras flutuantes libertadas sobem lentamente para a superfície com os ovos, permitindo, após a eclosão, que as larvas pelágicas flutuem e sejam livremente arrastadas pelas correntes até atingirem tamanho suficiente para deixar a superfície, migrando os juvenis para áreas mais profundas quando adultos.[38]
Na subordem Ceratioidei, a mais diversa dentro de Lophiiformes, os seus representantes apresentam elevado dimorfismo sexual, sendo que nos casos mais extremos, as fêmeas podem ter mais de 60 vezes o comprimento e ser cerca de meio milhão de vezes mais pesadas que os machos. Em algumas famílias, o macho, que é extremamente pequeno, pois no estado adulto atinge apenas cerca de 6–10 mm, procura ativamente por uma grande fêmea até se fixar sobre o seu corpo, permanente ou temporariamente, passando a alimentar-se do seu sangue. Esse hábito de vida, designado por simbiose sexual (ou parasitismo sexual segundo alguns autores), foi observado nas famílias Caulophrynidae, Neoceratiidae, Oneirodidae, Centrophrynidae e Linophrynidae.
Durante a metamorfose para a fase parasitária, os dentes dos machos são perdidos e substituídos por dentículos com formato de pinça em suas pontas, permitindo agarrar uma possível parceira. Em alguns táxons, ao prender-se na fêmea, o macho passa por uma fusão de tecidos, que é seguido da união dos sistemas circulatórios. Desse modo, o macho torna-se integralmente dependente da fêmea, sustentando-se dos nutrientes transportados pelos vasos sanguíneos desta, enquanto a fêmea hospedeira é capaz de se "auto-fertilizar", tornando-se um indivíduo semelhante a um hermafrodita típico.
Nestes casos de simbiose, os machos exibem uma imagem de como se fossem "absorvidos", onde a cabeça, desde o maxilar inferior até a parte posterior do crânio, é fundida à epiderme da fêmea. Geralmente, a união entre machos e fêmeas ocorre apenas com um indivíduo de cada sexo, contudo, múltiplas associações de machos em uma fêmea são comuns em alguns grupos.[5][6] Fêmeas não parasitadas não desenvolvem ovários, enquanto machos de vida livre não desenvolvem testículos e não apresentam um crescimento pós-metamórfico.[6]
Investigação bioquímica revelou que algumas das espécies de Lophiiformes que utilizam ligação permanente entre macho e fêmea carecem em grande parte dos genes que codificam as moléculas de complexo principal de histocompatibilidade (MHC), como se tivessem eliminado a capacidade de reconhecimento imunológico em favor da fusão de tecidos. Essa ausência de parte do MHC inibe a função das células T, que têm por função eliminar células infectadas ou atacar tecidos estranhos, dando origem ao processo de rejeição do órgão. Uma análise posterior indicou que os anticorpos estão ausentes em algumas espécies.[39]
Relevância económica e conservação
[editar | editar código-fonte]Diversas espécies desta ordem são exploradas pela pesca comercial e em várias regiões. A espécie Lophius gastrophysus, popularmente conhecida como tamboril-pescador, é considerada uma iguaria na Europa, Japão e América do Norte, sendo atualmente uma das pescarias mais valiosas do litoral brasileiro, com capturas usadas principalmente para exportação.[20][38]
As principal ameaça que afecta os membros desta ordem é a perda de habitat. A urbanização costeira em especial resulta em impactos locais para muitas espécies marinhas por meio da modificação do habitat, introdução de espécies invasoras e poluição. Espécies com baixa fecundidade, dispersão restrita e pequenas faixas geográficas podem ser mais suscetíveis a esses impactos, causando declínios populacionais.
Isso se aplica por exemplo para a espécie Brachionichthys hirsutus da família Brachionichthyidae, uma família de Lophiiformes endémica das águas australianas. A limitada capacidade de dispersão sugere que B. hirsutus pode depender de habitats específicos e espacialmente restritos e, portanto, pode ser particularmente suscetível à degradação do habitat. Esta degradação de habitat neste caso resulta da urbanização costeira, introdução de estrelas do mar do Pacífico Norte (Asterias amurensis), distúrbios pelo postos de atracação e mudanças climáticas.[40]
Um outro fator é a poluição dos oceanos por exemplo pelas emissões submarinas de esgoto ou dragagens de sedimento. Isto pode afetar as espécies de Lophiiformes direta ou indiretamente. Na ilha Bare, em Botany Bay (Austrália), ocorre uma espécie de Lophiiformes ("Bare Island anglerfish" ) que depende das esponjas existentes nessa região para se esconder e forragear. Como essas esponjas são sensíveis à poluição das águas, acabam morrendo e a sua ausência resulta na perda de habitat para a espécie.[41]
Referências
[editar | editar código-fonte]- ↑ Infopédia. «lofiiforme | Dicionário Infopédia da Língua Portuguesa». infopedia.pt - Porto Editora. Consultado em 28 de outubro de 2022
- ↑ Patterson, C. & Rosen, D.E. 1989. The Paracanthopterygii revisited: Order and disorder. In D.M. Cohen (editor), Papers on the Systematics of Gadiform Fishes. Science Series, 32:5-36.
- ↑ a b c d e Shedlock, Andrew M. (2003). «Molecular systematics and life history evolution of anglerfishes (Teleostei: Lophiiformes): Evidence from mitochondrial DNA». Steenstrupia
- ↑ Masaki, Miya et. al. (2010). «Evolutionary history of anglerfishes (Teleostei: Lophiiformes): a mitogenomic perspective». BMC Evolutionary Biology
- ↑ a b W. Pietsch, Theodore (2005). «Dimorphism, parasitism, and sex revisited: modes of reproduction among deep-sea ceratioid anglerfishes (Teleostei: Lophiiformes)». Ichthyological Research
- ↑ a b c d e f g h i j k l m n o p q r s Nelson, Grande & Wilson, Joseph S., Terry C. & Mark V. (2016). Fishes Of the World. New Jersey: John Wiley & Sons,
- ↑ Froese, Rainer, and Daniel Pauly, eds. (2006). Espécies de Lophiiformes em FishBase. Versão de Fevereiro de 2006.
- ↑ Freed, Lindsay L; Easson, Cole; Baker, Lydia J; Fenolio, Danté; Sutton, Tracey T; Khan, Yasmin; Blackwelder, Patricia; Hendry, Tory A; Lopez, Jose V (1 de outubro de 2019). «Characterization of the microbiome and bioluminescent symbionts across life stages of Ceratioid Anglerfishes of the Gulf of Mexico». FEMS Microbiology Ecology (em inglês). 95 (10): fiz146. ISSN 0168-6496. PMC 6778416
 . PMID 31504465. doi:10.1093/femsec/fiz146
. PMID 31504465. doi:10.1093/femsec/fiz146
- ↑ Baker, Lydia J; Freed, Lindsay L; Easson, Cole G; Lopez, Jose V; Fenolio, Danté; Sutton, Tracey T; Nyholm, Spencer V; Hendry, Tory A (1 de outubro de 2019). «Diverse deep-sea anglerfishes share a genetically reduced luminous symbiont that is acquired from the environment». eLife (em inglês). 8: e47606. ISSN 2050-084X. PMC 6773444. PMID 31571583. doi:10.7554/eLife.47606
- ↑ Pietsch, Theodore W. (25 de agosto de 2005). «Dimorphism, parasitism, and sex revisited: modes of reproduction among deep-sea ceratioid anglerfishes (Teleostei: Lophiiformes)». Ichthyological Research. 52 (3): 207–236. doi:10.1007/s10228-005-0286-2
- ↑ «Anglerfish». National Geographic. 10 de setembro de 2010. Consultado em 28 de fevereiro de 2019
- ↑ «Fish Identification». fishbase.org. Consultado em 28 de janeiro de 2019
- ↑ «Camouflage». Consultado em 22 de janeiro de 2018
- ↑ Nelson, Hans-Peter Schultze & Wilson, Joseph S., Hans-Peter & Mark V. H. (2010). Origin and Phylogenetic Interrelationships of Teleosts. München: Verlag Dr. Friedrich Pfeil
- ↑ Este artigo incorpora texto (em inglês) da Encyclopædia Britannica (11.ª edição), publicação em domínio público.
- ↑ «Anglerfish». National Geographic. 10 de setembro de 2010. Consultado em 18 de abril de 2018
- ↑ O'Day, William T. (1974). Bacterial Luminescence in the Deep-Sea Anglerfish (PDF). LA: Natural History Museum of Los Angeles County
- ↑ a b Munk, Ole; Hansen, Kjeld; Herring, Peter J. (2009). «On the Development and Structure of the Escal Light Organ of Some Melanocetid Deep Sea Anglerfishes (Pisces: Ceratioidei)». Journal of the Marine Biological Association of the United Kingdom. 78 (4). 1321 páginas. ISSN 0025-3154. doi:10.1017/S0025315400044520
- ↑ Haygood, M.G. 1993. Light organ symbioses in fishes. Crit. Rev. Microbiol. 19: 191-216.
- ↑ a b Guimarães de Abreu, Marta et. al. (2008). «Caracterização sensorial e análise bacteriológica do peixe-sapo (Lophius gastrophysus) refrigerado e irradiado». Ciência Rural
- ↑ Haygood, Margo G.; Distel, Daniel L. (Maio de 1993). «Bioluminescent symbionts of flashlight fishes and deep-sea anglerfishes form unique lineages related to the genus Vibrio». Nature (em inglês). 363 (6425): 154–156. ISSN 0028-0836. doi:10.1038/363154a0
- ↑ Hendry, Tory A.; Freed, Lindsay L.; Fader, Dana; Fenolio, Danté; Sutton, Tracey T.; Lopez, Jose V. (26 de junho de 2018). Moran, Nancy A., ed. «Ongoing Transposon-Mediated Genome Reduction in the Luminous Bacterial Symbionts of Deep-Sea Ceratioid Anglerfishes». mBio (em inglês). 9 (3): e01033–18, /mbio/9/3/mBio.01033–18.atom. ISSN 2150-7511. PMC 6020299. PMID 29946051. doi:10.1128/mBio.01033-18
- ↑ Miya, M.; T. Pietsch; J. Orr; R. Arnold; T. Satoh; A. Shedlock; H. Ho; M. Shimazaki; M. Yabe (2010). «Evolutionary history of anglerfishes (Teleostei: Lophiiformes): a mitogenomic perspective». BMC Evolutionary Biology. 10: 58. PMC 2836326. PMID 20178642. doi:10.1186/1471-2148-10-58
- ↑ Sepkoski, John J. (2002). «A compendium of fossil marine animal genera». Bulletins of American paleontology
- ↑ Pough, Janis & Heiser, F. Harvey, Christine M. & John B. (2006). Vertebrate Life. [S.l.]: Pearson Education
- ↑ Miya, Pietsch, et al.: Evolutionary history of anglerfishes (Teleostei: Lophiiformes): a mitogenomic perspective. BMC Evolutionary Biology, 10 (1), p. 58, Feb 2010 doi:10.1186/1471-2148-10-58
- ↑ Thomas J. Near, Ron I. Eytan, Alex Dornburg, Kristen L. Kuhn, Jon A. Moore, Matthew P. Davis, Peter C. Wainwright, Matt Friedman & W. Leo Smith: Resolution of ray-finned fish phylogeny and timing of diversification. PNAS, 2012, doi:10.1073/pnas.1206625109
- ↑ E. O. Wiley & G. David Johnson: A teleost classification based on monophyletic groups. pp. 123–182 in Joseph S. Nelson, Hans-Peter Schultze & Mark V. H. Wilson: Origin and Phylogenetic Interrelationships of Teleosts. 2010, Verlag Dr. Friedrich Pfeil, München, ISBN 978-3-89937-107-9.
- ↑ Theodore W. Pietsch, Lophiiformes (Anglerfishes).
- ↑ Ricardo Betancur-R, Edward O. Wiley, Gloria Arratia, Arturo Acero, Nicolas Bailly, Masaki Miya, Guillaume Lecointre und Guillermo Ortí: Phylogenetic classification of bony fishes. BMC Evolutionary Biology, BMC series – Juli 2017, DOI: 10.1186/s12862-017-0958-3
- ↑ Valentim, Maria de Fátima Moraes; Erica Pellegrini Caramaschi & Marcelo Vianna (2007) "Biologia e ecologia de peixes do género Lophius (Lophiidae, Lophiiformes), com ênfase em Lophius gastrophysus Miranda-Ribeiro, 1915: status atual." Oecologia Brasiliensis, ISSN 1981-9366, Vol. 11, Nº. 4, 2007 , pags. 503-520
- ↑ Fishbase: Antennarius biocellatus.
- ↑ «ScienceDirect». www.sciencedirect.com. doi:10.1016/0967-0637(95)00061-a. Consultado em 7 de maio de 2019
- ↑ a b Yasugi, Masaki; Hori, Michio (2016). «Predominance of parallel- and cross-predation in anglerfish». Marine Ecology (em inglês). 37 (3): 576–587. ISSN 1439-0485. doi:10.1111/maec.12309
- ↑ a b c d Karplus, Ilan (7 de abril de 2014). Symbiosis in Fishes: The Biology of Interspecific Partnerships (em inglês). Oxford: John Wiley & Sons. ISBN 9781118759769. doi:10.1002/9781118759769
- ↑ Smith, W. Leo; Sparks, John S.; Davis, Matthew P. (8 de junho de 2016). «Repeated and Widespread Evolution of Bioluminescence in Marine Fishes». PLOS ONE (em inglês). 11 (6): e0155154. ISSN 1932-6203. PMC 4898709. PMID 27276229. doi:10.1371/journal.pone.0155154
- ↑ «ScienceDirect». www.sciencedirect.com. Consultado em 7 de maio de 2019
- ↑ a b Valentim, Maria de Fátima Moraes; Caramaschi, Érica Pellegrini; Viana, Marcelo (dezembro de 2007). «Biologia e ecologia de peixes do género Lophius (Lophiidae, Lophiiformes) com ênfase em Lophius gastrophysus Miranda-Ribeiro, 1915: status atual». Oecologia Brasiliensis. 11 (04): 503–520. ISSN 1980-6442. doi:10.4257/oeco.2007.1104.04
- ↑ Malewar, Amit (2 de agosto de 2020). «Deep-sea anglerfishes sacrifice their immune system while mating». Tech Explorist (em inglês). Consultado em 6 de agosto de 2020
- ↑ Flynn, David J. H.; Green, Mark A.; Wright, Jeffrey T.; Barrett, Neville S.; Lynch, Tim P.; Wong, Lincoln S. C. (13 de agosto de 2018). «Local densities and habitat preference of the critically endangered spotted handfish (Brachionichthys hirsutus): Large scale field trial of GPS parameterised underwater visual census and diver attached camera». PLOS ONE (em inglês). 13 (8): e0201518. ISSN 1932-6203. doi:10.1371/journal.pone.0201518
- ↑ Arnold, Rachel (Janeiro de 2011). «In With the New: A New Anglerfish Species is Described in Botany Bay» (PDF). Consultado em 13 de junho de 2019
Galeria
[editar | editar código-fonte]-

-
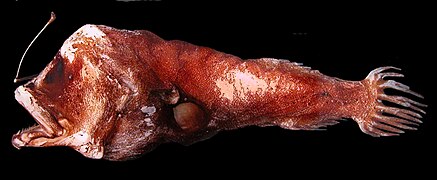
-

-

-

-
 Gigantactis sp. (Gigantactinidae).
Gigantactis sp. (Gigantactinidae). -

-

-

-

-

-









