
Cérebro
| Cérebro | |
|---|---|
 Cérebro de um chimpanzé | |
| Identificadores | |
| Latim | encephalon |
| MeSH | D001921 |
O cérebro é o principal órgão e centro do sistema nervoso em todos os animais vertebrados, e em muitos invertebrados. Alguns animais primitivos como os celenterados (água-viva e pólipo) e equinodermas (estrela-do-mar) possuem um sistema nervoso descentralizado sem cérebro, enquanto as esponjas-do-mar não possuem sistema nervoso. Nos vertebrados o cérebro localiza-se na cabeça protegido pelo crânio, próximo aos aparatos sensoriais primários: visão, audição, equilíbrio, tato, paladar e olfato. Num senso estrito, cérebro é o conjunto das estruturas nervosas derivadas do prosencéfalo (diencéfalo e telencéfalo). Em linguagem corrente, este termo pode designar o encéfalo como um todo, o telencéfalo ou o córtex cerebral.
Os cérebros podem ser extremamente complexos. O cérebro humano contém cerca de 86 bilhões de neurônios,[1] ligados por mais de 1 000 conexões sinápticas cada. Esses neurônios comunicam-se por meio de fibras protoplasmáticas chamadas axônio, que conduzem pulsos em sinais chamados potencial de ação para partes distantes do cérebro e do corpo e as encaminham para serem recebidas por células específicas.
De um ponto de vista filosófico, pode-se dizer que a função mais importante do cérebro é servir como estrutura física subjacente da mente. Do ponto de vista biológico, entretanto, a função mais importante do cérebro é a de gerador de comportamentos que promovam o bem-estar de um animal. O cérebro controla o comportamento, seja ativando músculos, seja causando a secreção de substâncias químicas, como os hormônios. Nem todos os comportamentos precisam de um cérebro. Mesmo organismos unicelulares são capazes de extrair informação do ambiente e responderem de acordo.[2] As esponjas, às quais falta um sistema nervoso central, são capazes de coordenar suas contrações corporais, e até mesmo de se locomoverem.[3] Nos vertebrados, a própria coluna vertebral contém circuitos neurais capazes de gerar respostas reflexas, assim como padrões motores simples, como nadar ou andar.[4] Entretanto, o controle sofisticado do comportamento, baseado em um sistema sensorial complexo requer a capacidade de integração de informações de um cérebro centralizado.
Apesar do rápido avanço científico, muito do funcionamento do cérebro continua um mistério. As operações individuais de neurônios e sinapses hoje são compreendidas com detalhamento considerável, mas o modo como eles cooperam em grupos de milhares ou milhões tem sido difícil de decifrar. Métodos de observação como registros de EEG e imageamento funcional cerebral mostram que as operações cerebrais são altamente organizadas, mas estes métodos não têm resolução suficiente para revelar a atividade de neurônios individualmente. Assim, mesmo os princípios mais fundamentais das redes de computação neural podem ficar, em grande medida, a serem descobertos por futuros pesquisadores.[5] O cérebro tem duas partes que ajudam no desenvolvimento: cerebelo e encéfalo.
Desenvolvimento
[editar | editar código-fonte]
O encéfalo não apenas cresce, ele se desenvolve em uma sequência muito bem orquestrada,[6] muitos neurônios são criados em zonas especiais que contêm células-tronco, e então migram pelo tecido para chegarem a sua localização final.[6] No córtex, por exemplo, o primeiro estágio de desenvolvimento é a formação de uma "plataforma" por um grupo especial de células gliais, chamadas glia radiais , que projetam fibras verticalmente através do córtex. Os neurônios corticais novos são criados na base do córtex, então "escalam" estas fibras radiais até chegarem às camadas que estão destinados a ocupar enquanto adultos.
Uma vez em seu lugar, o neurônio começa a estender dendritos e um axônio a seu redor.[6] Os axônios, por geralmente se estenderem a grande distância do corpo celular e terem de fazer contato com alvos específicos, crescem de modo particularmente complexo. A ponta de um axônio em crescimento consiste de uma bolha de protoplasma chamada "cone de crescimento", repleta de receptores químicos. Estes receptores sentem o ambiente local, fazendo o cone de crescimento ser atraído ou repelido por vários elementos celulares, sendo atraído a uma direção em particular em cada ponto de seu trajeto. O resultado deste processo de direcionamento é que o cone de crescimento navega através do cérebro até atingir sua área de destino, onde outros indicadores químicos o fazem iniciar a formação de sinapses. Levando em conta todo o encéfalo, muitos milhares de genes dão origem a proteínas que influenciam o direcionamento do axônio.


Entretanto, a rede sináptica que se forma é apenas parcialmente determinada pelos genes. Em muitas partes do encéfalo, há inicialmente um "supercrescimento" de axônios, que então são "ceifados" por mecanismos que dependem da atividade neural.[6] Na projeção do olho para o mesencéfalo, por exemplo, a estrutura adulta apresenta uma organização muito precisa, conectando cada ponto da superfície da retina a um ponto correspondente numa camada mesencefálica. Nos primeiros estágios de desenvolvimento, cada axônio da retina é guiado para a região correta do mesencéfalo por indicadores químicos, mas então se ramifica profusamente e faz contato inicial com um amplo feixe de neurônios do mesencéfalo. A retina, antes do nascimento, possui mecanismos especiais que a fazem gerar ondas de atividade que se originam em algum ponto e se propagam lentamente pela superfície retinal.[8] Estas ondas são úteis por ativarem ao mesmo tempo os neurônios vizinhos: quer dizer, elas produzem um padrão de atividade neural que contém informação sobre o arranjo espacial dos neurônios. Esta informação é utilizada no mesencéfalo por um mecanismo que faz as sinapses enfraquecerem, e finalmente desaparecerem, se a atividade em um axônio não for seguida pela ativação da célula-alvo. O resultado deste processo sofisticado é a gradual afinação e consolidação do sistema, até adquirir a forma final adulta. Processos semelhantes têm lugar em outras áreas do cérebro: uma matriz sináptica inicial é gerada, resultado do direcionamento químico geneticamente determinado, mas então é gradualmente refinada por mecanismos dependentes da atividade, parte controlados pela dinâmica interna, e parte por estímulos sensórios externos. Em alguns casos, assim como no sistema retina-mesencéfalo, os padrões de atividade dependem de mecanismos que operam apenas no cérebro em desenvolvimento, e aparentemente existem somente com o fim de guiar o desenvolvimento.
No ser humano e em muitos outros mamíferos, novos neurônios são criados principalmente antes do nascimento, e o cérebro infantil contém número significativamente maior do que o adulto.[6] Há entretanto umas poucas áreas onde novos neurônios continuam a ser criados durante a vida. As duas áreas para as quais o fato é pacífico são o bulbo olfatório e o giro dentado do hipocampo, onde há evidências de que novos neurônios estão envolvidos no armazenamento de memórias recentes. Com estas exceções, entretanto, o conjunto dos neurônios que estão presentes na primeira infância é o mesmo para o resto da vida. (Células gliais são diferentes: assim como a maioria dos tipos de células do corpo, estas se reproduzem ao longo da vida.) Apesar de o conjunto de neurônios já estar praticamente todo no lugar quando do nascimento, suas conexões axonais continuam a se desenvolver ainda por longo tempo. No ser humano, a mielinização não está completada até a adolescência.[6]
Houve longo debate sobre se as características da mente, personalidade e inteligência podem ser atribuídas à hereditariedade ou à criação; o debate "inato ou adquirido".[6] Não é uma questão apenas filosófica: ela assume grande relevância prática para pais e educadores. Apesar de muitos detalhes ainda precisarem ser esclarecidos, a neurociência mostra claramente que ambos fatores são essenciais. Os genes determinam a forma geral do encéfalo, e determinam como o encéfalo reage à experiência. A experiência, entretanto, é necessária para refinar a matriz de conexões sinápticas. Em alguns aspectos, esta (a matriz) é em grande parte uma questão de presença ou ausência de experiência durante períodos críticos de desenvolvimento.[6] Em outros aspectos, a quantidade e a qualidade da experiência pode ser mais relevante: por exemplo, há evidências substanciais de que animais criados em ambientes ricos (riqueza de estímulos) têm córtex mais espesso do que animais cujos níveis de estimulação são restritos.[6]
Estrutura macroscópica
[editar | editar código-fonte]
1. Encéfalo frontal
2. Telencéfalo
3. Diencéfalo
4. Tronco cerebral
5. Mesencéfalo
6. Ponte
7. Bulbo raquidiano
8. Cerebelo
9. Medula espinhal
O cérebro é a mais complexa estrutura biológica conhecida,[9] e compará-lo entre diferentes espécies mesmo nos aspectos básicos não é uma tarefa fácil. Porém, existem princípios comuns na arquitetura cerebral que se aplicam a uma vasta gama de espécies, que são revelados principalmente por três abordagens:
- A evolutiva que compara estruturas cerebrais de diferentes espécies e utiliza o princípio de que recursos encontrados em um determinado ramo também estavam presentes em seus ancestrais.
- A abordagem desenvolvimentista analisa como a forma do cérebro se desenvolve desde a fase embrionária até a fase adulta.
- A abordagem genética analisa expressão gênica em diversas partes do cérebro em toda uma gama de espécies. Cada abordagem complementa e informa as outras duas.
O córtex cerebral é a parte do cérebro que melhor distingue os mamíferos dos outros vertebrados, primatas de outros mamíferos e humanos de outros primatas. Em vertebrados não mamíferos, a superfície do telencéfalo é forrada por uma estrutura em camadas relativamente simples chamada pallium.[10] Nos mamíferos, o pallium é envolvido em uma estrutura de 6 camadas chamada neocortex. Em primatas o neocortex é mais avantajado em comparação aos não primatas, especialmente a parte chamada lobo frontal. Nos seres humanos, este alargamento dos lobos frontais é levado de uma extremidade à outra, e de outras partes do córtex também se tornam bastante grandes e complexas. A relação entre tamanho cerebral, tamanho corporal e outras variáveis são estudadas entre uma grande gama de espécies.
O tamanho do cérebro aumenta com o tamanho do corpo mas não proporcionalmente. A média em todas as ordens de mamíferos segue a lei de potência, com o expoente de cerca de 0,75.[11] Esta fórmula pode aplicar-se ao cérebro de um mamífero médio, mas cada família desvia do padrão, refletindo o nível de sofisticação em seu comportamento.[12] Por exemplo, os primatas têm cérebros de 5 a 10 vezes maior que o indicado pela fórmula. Predadores tendem a ter cérebros maiores. Quando aumenta o tamanho do cérebro de um mamífero, nem todas as partes aumentam na mesma proporção. Quanto maior o cérebro de uma espécie, maior a porção representada pelo córtex.[13]
Bilatérios
[editar | editar código-fonte]
Com exceção de umas poucas formas primitivas como as esponjas e águas-vivas, todos os animais existentes são bilaterais, ou seja, animais cujo corpo apresenta simetria bilateral (isto é, o lado direito e o esquerdo são imagens espelhadas um do outro).
Imagina-se que todos os bilatérios descendam de um ancestral comum, surgido no início do período Cambriano, num período datado entre 550 e 600 milhões de anos antes do presente.[14] Este ancestral tinha a forma de um simples verme tubular de corpo segmentado, e num nível abstrato, este formato de verme continua presente no esquema dos corpos e sistemas nervosos de todos os bilatérios modernos, inclusive o ser humano.[15] A forma geral de corpo bilatério é a de um tubo com uma cavidade digestiva oca indo da boca ao ânus, e um cordão neural com um alargamento (um gânglio) para cada segmento corporal, com um gânglio excepcionalmente grande na frente, chamado de "cérebro".
Invertebrados
[editar | editar código-fonte]Em muitos invertebrados - insetos, moluscos, vermes de vários tipos etc. - os componentes do cérebro e a sua organização difere tanto do padrão dos vertebrados que se torna difícil fazer comparações com algum significado, exceto com base na genética. Dois grupos de invertebrados possuem cérebros notavelmente complexos: artrópodes (insetos, crustáceos, aracnídeos, e outros) e cefalópodes (polvos, lulas e moluscos semelhantes).[16] Os cérebros dos artrópodes e cefalópodes chegam de dois cordões neurais paralelos que se estendem pelo corpo do animal. Artrópodes possuem um cérebro central com três divisões e grandes lobos ópticos atrás de cada olho, para processamento visual.[16] Cefalópodes têm os maiores cérebros entre os invertebrados. O cérebro do polvo, em particular, é altamente desenvolvido, comparável em complexidade com os cérebros de alguns vertebrados.
Somente uns poucos invertebrados tiveram seus cérebros estudados intensivamente. A grande lesma-do-mar Aplysia foi escolhida pelo prêmio Nobel de neurofisiologia Eric Kandel, pela simplicidade e acessibilidade de seu sistema nervoso, como modelo para o estudo das bases celulares do aprendizado e memória, e submetida a centenas de experimentos.[17] Os cérebros invertebrados mais amplamente estudados, entretanto, pertencem à mosca-da-fruta Drosophila e à pequena nematoda Caenorhabditis elegans.
.jpg)
Pela abundância de técnicas disponíveis para estudar sua genética, a mosca-da-fruta tornou-se o objeto natural no estudo do papel dos genes no desenvolvimento do cérebro.[18] Notavelmente, muitos aspectos neurogenéticos da Drosophila mostraram-se relevantes para os humanos: os primeiros genes do relógio biológico, por exemplo, foram identificados ao se examinar Drosophilae mutantes que apresentavam ciclos irregulares na atividade diária.[19] Uma pesquisa nos genomas dos vertebrados descobriu um conjunto de genes análogos que desempenham papel similar no relógio biológico de camundongos - e portanto, quase que certamente no relógio biológico humano.[20]
Como a Drosophila, a C. elegans foi estudada amplamente por sua importância para a genética.[21] No início dos anos 1970, Sydney Brenner a escolheu como organismos modelo para estudar o modo como os genes controlam o desenvolvimento. Uma das vantagens de trabalhar com este verme é que a estrutura corporal é bastante estereotipado: o sistema nervoso da forma hermafrodita possui exatamente 302 neurônios, sempre nos mesmos lugares, formando conexões sinápticas idênticas em cada verme.[22] Num projeto heroico, a equipe de Brenner fatiou vermes em milhares de seções ultrafinas e fotografou cada seção num microscópio eletrônico, então encaixou visualmente as fibras de seção para seção, a fim de mapear cada neurônio e cada sinapse de todo o corpo.[23] Nada que se aproxime deste nível de detalhe está disponível para outro organismo, e a informação obtida permitiu uma multitude de estudos que não teria sido possível de outro modo.
Vertebrados
[editar | editar código-fonte]
Os cérebros dos vertebrados são feitos de um tecido muito mole, de textura comparável à da geleia.[6] Quando vivo, o tecido cerebral é rosado por fora e branco por dentro, com pequenas variações de cor. Nos vertebrados, o cérebro é circundado por um sistema de membranas de tecido conjuntivo chamadas meninges, que o separam do crânio.[24] Esta cobertura em três camadas é composta (de fora para dentro) pela dura-máter (matéria dura), aracnoide-máter (matéria-aranha), e pia-máter (matéria macia). A aracnoide e a pia são fisicamente conectadas, e frequentemente consideradas uma única camada, a pia-aracnoide. Sob a aracnoide fica o espaço sub-aracnoide, que contém fluido cérebro-espinal (FCE), que circula pelos pequenos espaços intercelulares e por cavidades chamadas ventrículos, e serve para nutrir, sustentar e proteger o tecido cerebral. Vaso sanguíneo entram no sistema nervoso central pelo espaço perivascular acima da pia-máter. As células das paredes destes vasos são firmemente unidas, formando a barreira sangue-cérebro, que protege o cérebro de toxinas que possam entrar pelo sangue.
Os primeiros vertebrados apareceram há mais de 500 milhões de anos (Ma), durante o período Cambriano, e talvez lembrassem uma enguia.[25] Os tubarões apareceram por volta de 450 Ma, anfíbios 400 Ma, répteis por volta de 350 Ma e mamíferos uns 200 Ma. Não seria correto dizer que qualquer espécie atual é mais primitiva do que outra, já que todas têm sua história evolutiva igualmente longas, mas os cérebros dos modernos peixe-bruxa, lampreias, tubarões, anfíbios, répteis e mamíferos apresentam uma gradação de tamanho e complexidade que, grosso modo, segue a sequência evolutiva.[26] Todos estes cérebros contêm basicamente o mesmo conjunto de elementos anatômicos, mas muitos destes são rudimentares, como no peixe-bruxa, enquanto nos mamíferos as partes frontais são altamente elaboradas e expandidas.

Todos os cérebros vertebrados partilham de uma mesma forma fundamental, que pode ser apreciada mais facilmente examinando como eles se desenvolvem.[6] O sistema nervoso aparece na forma de uma fina tira de tecido que corre pelo dorso do embrião. Esta tira engrossa e então se dobra para formar um tubo oco. A extremidade frontal do tubo se desenvolve e forma o cérebro. Em sua forma mais recente, o cérebro aparece como três protuberâncias, que finalmente formarão o posencéfalo, o mesencéfalo e o rombencéfalo. Em muitas classes de vertebrados, o tamanho destas três partes permanece similar no adulto, mas nos mamíferos o posencéfalo fica muito maior que as outras partes, e o mesencéfalo bem pequeno.
Geralmente, os neuroanatomistas dividem o cérebro em seis regiões principais: o telencéfalo (hemisférios cerebrais), o diencéfalo (tálamo e hipotálamo), mesencéfalo, cerebelo, ponte e medula.[6] Cada área destas, por sua vez, possui uma estrutura interna complexa. Algumas áreas, como o córtex e o cerebelo, constituem-se de camadas, dobradas ou enroladas para caberem no espaço disponível. Outras áreas são constituídas de aglomerados de numerosos pequenos núcleos. Se forem feitas distinções estritas baseadas na estrutura neural, química e conectividade, milhares de áreas diferentes podem ser identificadas no cérebro dos vertebrados.
Alguns ramos de evolução dos vertebrados levaram a mudanças substanciais no formato cerebral, especialmente no posencéfalo. O cérebro do tubarão apresenta os elementos básicos numa disposição simples, mas nos peixes teleósteos (grande maioria das espécies modernas), o posencéfalo tornou-se revirado, como uma meia virada do avesso. Nas aves, também, há grandes mudanças no formato.[27] Por muito tempo, se pensou que uma das principais estruturas do posencéfalo das aves, a espinha dorsal ventricular, correspondesse ao gânglio basal dos mamíferos, mas hoje acredita-se estar mais relacionado ao neocórtex.[28]
Diversas áreas cerebrais mantêm a mesma identidade entre todos os vertebrados, do peixes-bruxa ao ser humano. Segue uma lista de algumas das áreas mais importantes, com breve descrição de suas funções como são entendidas atualmente (mas note-se que ainda existe algum grau de discordância a respeito das funções da maioria das áreas):
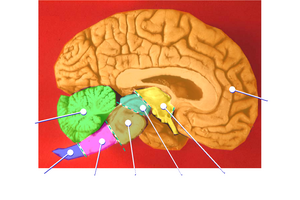
- A medula, ao longo do cordão espinhal, contém vários pequenos núcleos envolvidos numa ampla variedade de funções sensórias e motoras.
- O hipotálamo é uma pequena região na base do posencéfalo, cuja complexidade não corresponde ao tamanho. É composto de numerosos pequenos núcleos, cada um com conexões distintas e neuroquímica idem. O hipotálamo é a estação central de controle dos ciclos de sono/alerta, controle de fome e sede, controle da liberação de hormônios e muitas outras funções biológicas críticas.[29]
- Como o hipotálamo, o tálamo é um conjunto de núcleos com funções diversas. Alguns estão envolvidos em retransmitir informações dos e para os hemisférios cerebrais. Outros estão envolvidos na motivação. A região subtalâmica (zona incerta) parece conter sistemas geradores de ação para diversos tipos de comportamentos "consumatórios", incluindo comer, beber, defecação e cópula.[30]
- O cerebelo modula as informações de outros sistemas cerebrais para fazê-las mais precisas. A remoção do cerebelo não impede um animal de fazer nada em particular, mas deixa suas ações hesitantes e desajeitadas. Tal precisão não é inata, mas aprendida por tentativa e erro. Aprender a andar de bicicleta é exemplo de um tipo de plasticidade neural que acontece majoritariamente dentro do cerebelo.[6]
- O teto, também chamado de "teto óptico" permite direcionar ações a determinado ponto no espaço. Nos mamíferos, é chamado de colículo superior, e sua função mais bem estudada é a de direcionar os movimentos oculares. Mas também dirige o movimento de alcançar. O teto recebe fortes estímulos visuais, mas também estímulos de outros sentidos que são úteis ao direcionamento de ações, como estímulos auditivos em corujas, estímulos dos órgãos termo-sensíveis em cobras, etc. Em alguns quais peixes, o teto é a maior porção do cérebro.[31]
- O pálio é uma camada de matéria cinzenta que fica na superfície do posencéfalo. Nos répteis e mamíferos, ela é chamada de córtex. O pálio está relacionado a múltiplas funções, incluindo o olfato e a memória espacial. Nos mamíferos, em que o córtex domina o cérebro, ele assume funções de várias regiões subcorticais.[32]
- O hipocampo, estritamente falando, é encontrado apenas em mamíferos. No entanto, a região da qual ele deriva, o pallium medial, tem correspondentes em todos os vertebrados. Há evidências de que esta parte do cérebro está envolvida na memória espacial e navegação de aves, peixes, répteis e mamíferos.[33]
- Os gânglios basais são um grupo de estruturas interconectadas do posencéfalo, das quais nosso entendimento aumentou consideravelmente nos últimos anos. A função primária dos gânglios basais parece ser a de seleção de ação. Eles mandam sinais inibitórios para todas as partes do cérebro que possam gerar ações, e nas circunstâncias certas pode liberar a inibição, de modo que os sistemas de geração de ação executem suas ações. Recompensas e punições têm seus efeitos neurais mais importantes sobre os gânglios basais.[34]
- O bulbo olfativo é uma estrutura especial que processa os sinais sensórios olfativos e envia seus resultados para a parte olfativa do pálio. É um elemento significativo do cérebro de muitos vertebrados, mas é muito reduzido nos primatas.[35]
Mamíferos
[editar | editar código-fonte]O rombencéfalo e o mesencéfalo dos mamíferos são em geral similares aos de outros vertebrados, mas diferenças gritantes aparecem no prosencéfalo, que é não só muito aumentado, mas diferenciado em sua estrutura.[36] Nos mamíferos, a maior parte da superfície dos hemisférios cerebrais é coberta por um isocórtex de seis camadas, mais complexo que o pallium de três camadas visto na maioria dos vertebrados. O hipocampo dos mamíferos também apresenta uma estrutura diferente.
A história evolucionária destas características mamíferas, especialmente o córtex de seis camadas, é difícil de reconstituir.[10] Isto deve-se, principalmente, ao desconhecimento de um fóssil de transição. Os ancestrais dos mamíferos, chamados sinápsidas, separam-se dos ancestrais dos répteis modernos e aves há cerca de 350 milhões de anos. Entretanto, a ramificação mais recente que vingou entre os mamíferos foi a separação entre monotremados (ornitorrinco e équidna), marsupiais (gambá, canguru) e placentários (maioria dos mamíferos atuais), que aconteceu há cerca de 120 milhões de anos. Os cérebros dos monotremados e dos marsupiais são diferentes dos cérebros placentários em alguns aspectos, mas possuem as estruturas corticais e do hipocampo inteiramente mamíferas. Assim, estas estruturas devem ter evoluído há entre 350 e 120 milhões de anos, período que não deixou evidências senão fósseis, que não conservam tecidos moles como o cérebro.
Em 2018 cientistas reviveram os cérebros de cerca de 200 porcos e os mantiveram vivos fora de seus corpos, produzindo uma onda cerebral plana comparável a alguém em estado comatoso, por 36 horas.[37]
Primatas (incluindo humanos)
[editar | editar código-fonte]O cérebro primata contém a mesma estrutura que o cérebro de outros mamíferos, mas é consideravelmente maior em relação ao tamanho do corpo.[13] Esse aumento de tamanho relaciona-se principalmente com uma grande expansão do córtex, com destaque para as áreas relativas a visão e antecipação.[38] A rede de processamento visual dos primatas é muito complexa, incluindo pelo menos 30 áreas diferenciáveis, com uma desconcertante rede de interconexões. Esses fatos contribuem para que o processamento visual utilize quase metade do cérebro. A outra parte do cérebro que tem grande aumento é o córtex pré-frontal, cujas funções são difíceis de sumarizar sucintamente, mas relacionam-se com planejamento, memória de trabalho, motivação, atenção, e controle de funções.
Estrutura microscópica
[editar | editar código-fonte]
O cérebro é composto de duas grandes classes de células, neurônios e células gliais.[6] Neurônios recebem mais atenção, mas, na verdade, as células gliais são mais frequentes, formando uma proporção de pelo menos 10 para 1. Existem diversos tipos de células gliais, que realizam um grande número de funções importantes como: suporte estrutural, suporte metabólico, isolamento, e guia para o desenvolvimento. A característica que torna os neurônios tão importantes é a capacidade de enviar sinais uns para os outros através de longas distâncias, algo que não ocorre nas células gliais.[6] Eles enviam esses sinais através de um axônio, uma fina fibra protoplasmática que parte do corpo celular e projeta-se, normalmente com inúmeras ramificações, para outras áreas, às vezes perto, às vezes em partes distantes do cérebro ou do corpo. A extensão de um axônio pode ser extraordinária: por exemplo, se uma célula piramidal do neocórtex fosse aumentada até que o tamanho de seu corpo fica-se do tamanho de um corpo humano, seu axônio, igualmente aumentado, seria um cabo com algumas polegadas de diâmetro, estendendo-se por mais de um quilômetro. Esses axônios transmitem sinais na forma de impulsos eletroquímicos chamados potenciais de ação, que duram menos que um milésimo de segundo e viajam através do axônio numa velocidade de 1 a 100 metros por segundo. Alguns neurônios emitem potenciais de ação constantemente, 10 a 100 vezes por segundo, normalmente em padrões temporais irregulares; outros neurônios ficam em repouso a maior parte do tempo, mas ocasionalmente emitem uma rajada de potenciais de ação.
Axônios transmitem sinais para outros neurônios, ou para células não-neuronais, através de uma junção especializada chamada sinapse.[6] Um único axônio pode fazer diversas conexões sinápticas. Quando um potencial de ação, viajando através do axônio, chega à sinapse, faz com que um composto químico chamado de neurotransmissor seja liberado. O neurotransmissor liga-se a moléculas receptoras na membrana da célula alvo. Alguns tipos de receptores neuronais são excitatórios, ou seja, eles aumentam a frequência dos potenciais de ação na célula alvo; outros receptores são inibitórios, ou seja, eles diminuem a frequência dos potenciais de ação; outros tem efeitos efeitos modulatórios complexos na célula alvo. Na verdade, são os axônios que preenchem a maior parte do espaço do cérebro.[6] Normalmente, grandes grupos deles viajam juntos em aglomerados chamados tratos de fibras nervosas. Em muitos casos, cada axônio é envolto por uma grossa bainha de uma substância lipídica chamada Mielina, que serve para aumentar muito a velocidade de propagação do potencial de ação. Mielina tem a coloração branca, então partes do cérebro preenchidas exclusivamente por fibras nervosas aparecem como substância branca, por outro lado, a substância cinzenta marca as áreas com altas densidades de corpos celulares neuronais.
Funcionamento
[editar | editar código-fonte]
De uma perspectiva biológica, a função do cérebro é gerar comportamentos que promovam a aptidão genética de um animal.[39] Para fazê-lo, ele extrai informações relevantes dos órgãos sensíveis para refinar as ações do animal. Sinais sensórios podem estimular respostas imediatas, como quando o sistema olfatório de um veado detecta o odor de um lobo; podem modular o padrão de atividade em andamento, como os efeitos dos ciclos de claridade-escuridão sobre o estado de sono-vigília de um organismo; ou suas informações podem ser armazenadas, para o caso de relevância futura. O cérebro gerencia sua complexa tarefa orquestrando subsistemas funcionais, que podem ser categorizados de várias formas: anatomicamente, quimicamente e funcionalmente.
Apesar do cérebro representar apenas 2% da massa corporal, ele é responsável por 20% de consumo total de oxigênio do corpo humano.[40]
Sistemas de neurotransmissores
[editar | editar código-fonte]Com poucas exceções, cada neurônio do cérebro libera o mesmo neurotransmissor químico, ou conjunto de neurotransmissores, em todas as conexões sinápticas que faz com outros neurônio.[6] Ainda assim, a grande maioria das drogas psicoativas produz efeitos ao alterar sistemas neurotransmissores que não envolvem diretamente a transmissão glutamatérgica ou a GABAérgica.[41] Drogas como cafeína, nicotina, heroína, cocaína, Prozac, Thorazine, etc., atuam sobre outros neurotransimssores. Muitos destes outros transmissores vêm de neurônios localizados em partes específicas do cérebro. A serotonina, por exemplo - o alvo primário de drogas antidepressivas e muitos suplementos dietéticos - origina-se em uma pequena área do tronco encefálico chamada núcleo da rafe. A norepinefrina, relacionada ao estado de alerta, origina-se em uma pequena área próxima, chamada de cerúleo. A histamina, enquanto neurotransmissor, vem de uma pequena parte do hipotálamo chamada núcleo tuberomamilar (a histamina também possui funções fora do sistema nervoso central, mas a função neurotransmissora é que faz os anti-histamínicos terem efeito sedativo). Outros neurotransmissores como a acetilcolina e a dopamina têm múltiplas fontes no cérebro, mas que não estão tão ubiquamente distribuídas quanto as de glutamato e GABA.
Sistemas sensórios
[editar | editar código-fonte]Uma das funções primárias do cérebro é extrair informação biologicamente relevante de receptores sensoriais.[6] Mesmo no cérebro humano, os processos sensórios vão bem além dos clássicos cinco sentidos da visão, audição, paladar, tato e olfato: nossos cérebros recebem informações sobre temperatura, equilíbrio, posição dos membros e da composição química da corrente sanguínea, entre outras coisas. Todas estas informações são detectadas por sensores especializados que enviam sinais para o cérebro. Entre não-humanos, podem estar presentes sentidos adicionais, como os sensores de calor presentes nas fossetas loreais das serpentes; ou os sentidos "convencionais" podem ser usados de modos não-convencionais, como no "sonar" auditivo dos morcegos. Cada sistema sensório possui suas idiossincrasias, mas aqui estão alguns princípios que se aplicam à maioria deles, usando o sentido da audição para exemplos específicos:[6]
- Cada sistema começa com células especializadas de "recepção sensorial". Estas são neurônios, mas diferente da maioria deles, elas não são controladas por estímulos sinápticos de outros neurônios; em vez disso, elas são ativadas por receptores ligados à membrana que são sensíveis a alguma modalidade física, como luz, temperatura, ou estiramento físico. Os axônios dos receptores sensórios adentram a medula espinhal ou o cérebro. No sentido da audição, os receptores localizam-se no ouvido interno, na cóclea, e são ativados por vibração;
- Para a maioria dos sentidos, há um "núcleo primário", ou um conjunto de núcleos, localizado no tronco cerebral, que coleta sinais das células-receptoras. Para o sentido da audição, há o núcleo coclear;
- Em muitos casos, há áreas subcorticais secundárias que extraem algum tipo de informação especial. No sentido da audição, o complexo olivar superior e o colículo inferior estão envolvidos na comparação dos sinais dos dois ouvidos para extrair informação sobre a direção da fonte sonora, entre outras funções;
- Cada sistema sensório também tem uma parte especial do tálamo dedicada a si, que serve como um retransmissor para o cérebro. Para o sistema auditivo, este é o núcleo geniculado medial.
- Para cada sistema sensório, há uma área cortical "primária" que recebe estímulos diretos da área de retransmissão talâmica. Para o sistema auditivo esta esta é o córtex auditivo primário, localizado na parte superior do lobo temporal;
- Também há geralmente um conjunto de áreas sensórias corticais "de alto nível", que analisam o estímulo sensório de modo específico. Para o sistema auditivo, há áreas que analisam a qualidade do som, ritmo, e padrões de mudança temporal, entre outros aspectos;
- Finalmente, há áreas multimodais que combinam estímulos de diferentes modalidades sensoriais, por exemplo auditivas e visuais. Neste ponto, os sinais atingiram partes do cérebro que são mais bem-descritas como integradoras do que como sensórias.
Todas estas regras têm exceções, por exemplo: (1) No sentido do tato (que na verdade é um conjunto de pelo menos meia dúzia de sentidos mecânicos), os estímulos sensórios se encerram principalmente no cordão espinhal, em neurônios que se projetam para o tronco cerebral. Para o sentido do olfato, não há qualquer retransmissor no tálamo; em vez disso, os sinais seguem diretamente da região encefálica primária - o bulbo olfatório - para o córtex.[6]
Sistema motor
[editar | editar código-fonte]Sistemas motores são áreas do cérebro que estão mais ou menos envolvidas na produção de movimentos corporais, isto é, na ativação de músculos. Com a exceção dos músculos que controlam os olhos, todos os músculos voluntários[42] do corpo são diretamente inervados por neurônios motores no cordão espinhal, que por sua vez são o último caminho comum do sistema gerador de movimento.[6] Neurônios motores espinhais são controlados tanto por circuitos neurais intrínsecos ao cordão espinhal quanto por estímulos originados no cérebro. Os circuitos espinhais intrínsecos executam várias respostas reflexas, e também contêm geradores de padrões para movimentos ritmados, como andar ou nadar. As conexões descendentes do encéfalo permitem controle mais sofisticado.[6]
O cérebro contém algumas áreas que se projetam diretamente para o cordão espinhal.[6] No nível mais baixo estão a as áreas motoras na medula e na ponte. Num nível mais alto se encontram áreas no mesencéfalo, como o núcleo rubro, que é responsável pela coordenação dos movimentos de braços e pernas. Num nível ainda mais alto, está o córtex motor primário, uma tira de tecido localizada no limite posterior do lobo frontal. O córtex motor primário propaga-se para as regiões motoras subcorticais, mas também envia uma projeção maciça diretamente para o cordão espinhal, através do assim chamado trato piramidal. Esta projeção córtico-espinhal direta é responsável pelo controle voluntário dos detalhes finos dos movimentos.[6]
Outras áreas "secundárias" do cérebro relacionadas ao movimento não se projetam diretamente ao cordão espinhal, mas em vez disso agem sobre as áreas motoras primárias corticais ou subcorticais. Dentre as áreas secundárias mais importante se encontram o córtex premotor, os gânglios basais e o cerebelo:[6]
- O córtex premotor (que na verdade é um grande complexo de áreas) une-se ao córtex motor primário e projeta-se nele. Enquanto os elementos do córtex motor primário mapeiam áreas específicas do corpo, os elementos do córtex premotor estão mais envolvidos nos movimentos coordenados de diversas partes do corpo.
- Os gânglios basais são um conjunto de estruturas na base do prosencéfalo que projetam-se para várias outras áreas relacionadas ao movimento. Sua função tem sido difícil de compreender, mas uma das teorias mais aceitas atualmente é de que eles tenham parte crucial na seleção de ação. Na maior parte do tempo, eles refreiam ações, enviando sinais inibitórios constantes para os sistemas geradores de ação, mas nas circunstâncias corretas, eles cessam esta inibição e assim permitem a seus alvos tomarem o controle do comportamento.
- O cerebelo é uma estrutura bastante distinta presa ao fundo do cérebro. Ele não controla ou origina comportamentos, mas gera sinais corretivos para tornar os movimentos mais precisos. Pessoas com danos cerebelares não ficam paralisadas em nenhum aspecto, mas seus movimentos corporais tornam-se erráticos e descoordenados.
Além de tudo acima, o cérebro e o cordão espinhal contêm extensos circuitos para o controle do sistema nervoso autônomo, que funciona pela secreção de hormônios e pela modulação dos músculos lisos do intestino. O sistema nervoso autônomo afeta o ritmo cardíaco, a digestão, o ritmo respiratório, a salivação, a perspiração, a urinação e a excitação sexual - mas muitas de suas funções não estão sob controle voluntário.[6]
Sistema de alerta
[editar | editar código-fonte]Talvez o aspecto mais óbvio do comportamento de qualquer animal é o ciclo diário que compreende dormir e acordar. O estado de alerta e atenção também é modulado em uma escala de tempo delicada por uma rede extensa de áreas cerebrais.[6]
Um componente chave do sistema da alerta é o núcleo supraquiasmático, uma pequena porção do hipotálamo localizada diretamente acima do ponto em que os nervos ópticos dos dois olhos cruzam.[43] O núcleo supraquiasmático contém o relógio biológico central do corpo. Os neurônios de lá mostram níveis de atividade que sobem e declinam em um período de 24 horas, os ritmos circadianos: essas flutuações de atividade são direcionadas por mudanças rítmicas na expressão de um determinado grupo de "genes relógio". O núcleo supraquiasmático continua a altera-se de acordo com tempo mesmo se for retirado do cérebro e colocado numa bandeja com uma solução nutritiva morna, entretanto ele apenas recebe a informação dos nervos ópticos, através do trato retino hipotalâmico, que permite que o ciclo claro-escuro calibre o relógio.
O núcleo supraquiasmático projeta-se a um grupo de áreas no hipotálamo, tronco encefálico e cérebro médio que estão relacionadas com a implementação dos ciclos sono-vigília. Um componente importante do sistema é a chamada formação reticular, um grupo de "amontoados" neuronais espalhados difusamente através do núcleo do tronco encefálico.[6] Neurônios reticulares enviam sinais para o tálamo, que, por sua vez, envia sinais de controle do nível de atividade para todo o córtex. Danos a formação reticular pode provocar um estado permanente de coma. Sono envolve grandes mudanças na atividade cerebral.[6]
Efeitos de doenças e lesões
[editar | editar código-fonte]Apesar de ser protegido pelo crânio e pelas meninges, envolvido pelo líquido cefalorraquidiano, e isolado da corrente sangüínea pela barreira hematoencefálica, a natureza sensível do cérebro o faz vulnerável a inúmeras doenças e diversos tipos de lesões. Esses problemas manifestam-se de maneira diferenciada em humanos em relação a outras espécies, por isso uma visão geral da patologia cerebral e seu possível tratamento são abordados nos artigos sobre cérebro humano, lesão cerebral, e neurologia.
Ver também
[editar | editar código-fonte]- serotonina
- encéfalo
- sistema nervoso
- morte cerebral
- paralisia cerebral
- memória
- sistema límbico
- Disfunção do lobo frontal
Referências
- ↑ F.A.C. Azevedo e colaboradores (2009) Equal numbers of neuronal and non-neuronal cells make the human brain an isometrically scaled-up primate brain; Journal of Comparative Neurology. pp. 532-541.
- ↑ Gehring, Wj (2005) New Perspectives on Eye Development and the Evolution of Eyes and Photoreceptors: The Evolution of Eyes and Brain, J Heredity (acessado em 26-04-2008)
- ↑ Nickel,M; Vitello, M; Brümmer, F (2002). «Dynamics and cellular movements in the locomotion of the sponge Tethya wilhelma». Integr Comp Biol. 42. 1285 páginas
- ↑ Grillner, S; Hellgren J, Ménard A, Saitoh K, Wikström MA (2005). «Mechanisms for selection of basic motor programs—roles for the striatum and pallidum.». Trends Neurosci. 28: 364–70. PMID 15935487. doi:10.1016/j.tins.2005.05.004
- ↑ van Hemmen, JL; Sejnowski TJ (2005). 23 Problems in Systems Neuroscience. [S.l.]: Oxford University Press. ISBN 9780195148220
- ↑ a b c d e f g h i j k l m n o p q r s t u v w x y z aa ab ac ad Purves, Dale.; Lichtman, Jeff W. (1985). Principles of neural development. Sunderland, Mass.: Sinauer Associates. ISBN 978-0-87893-744-8. OCLC 10798963
- ↑ Kandel, Eric R.; Schwartz, James Harris; Jessell, Thomas M. (2000). Principles of neural science 4th ed. New York: McGraw-Hill. ISBN 978-0-8385-7701-1. OCLC 42073108
- ↑ Wong, 1999
- ↑ Shepherd GM (1994). Neurobiology. [S.l.]: Oxford University Press. ISBN 9780195088434
- ↑ a b Aboitiz, F; Morales D, Montiel J (2003). «The evolutionary origin of the mammalian isocortex: Towards an integrated developmental and functional approach.». Behav Brain Sci. 26: 535–52. PMID 15179935. doi:10.1017/S0140525X03000128. Consultado em 22 de dezembro de 2008. Arquivado do original em 29 de janeiro de 2009
- ↑ Armstrong, E (1983). «Relative brain size and metabolism in mammals.». Science. 220: 1302–4. PMID 6407108. doi:10.1126/science.6407108
- ↑ Jerison, HJ (1973). Evolution of the Brain and Intelligence. [S.l.]: Academic Press. pp. http://books.google.com/books?id=xTpDAAAACAAJ. ISBN 9780123852502
- ↑ a b Finlay, BL; Darlington RB, Nicastro N (2001). «Developmental structure in brain evolution.» (PDF). Behav Brain Sci. 20: 263–308. PMID 11530543. Consultado em 29 de dezembro de 2008. Arquivado do original (PDF) em 31 de outubro de 2008
- ↑ Balavoine, G; Adoutte A (2003). «The segmented Urbilateria: A testable scenario.». Int Comp Biology. 43: 137–47. doi:10.1093/icb/43.1.137
- ↑ Schmidt-Rhaesa, A (2007). The Evolution of Organ Systems. [S.l.]: Oxford University Press. ISBN 9780198566694
- ↑ a b Butler, AB (2000). «Chordate Evolution and the Origin of Craniates: An Old Brain in a New Head.». Anat Rec. 261: 111–25. PMID 10867629. doi:10.1002/1097-0185(20000615)261:3<111::AID-AR6>3.0.CO;2-F[ligação inativa]
- ↑ Kandel, ER (2007). In Search of Memory: The Emergence of a New Science of Mind. [S.l.]: WW Norton. pp. http://books.google.com/books?id=LURy5gojaDoC. ISBN 9780393329377
- ↑ «Flybrain: An online atlas and database of the drosophila nervous system». Consultado em 23 de dezembro de 2008. Arquivado do original em 16 de maio de 2016
- ↑ Konopka, RJ; Benzer S (1971). «Clock mutants of Drosophila melanogaster.». PNAS. 68: 2112–6. PMID 5002428. doi:10.1073/pnas.68.9.2112
- ↑ Shin, HS; Bargiello TA, Clark BT, Jackson FR, Young MW (1985). «An unusual coding sequence from a Drosophila clock gene is conserved in vertebrates.». Nature. 317: 445–8. PMID 2413365. doi:10.1038/317445a0
- ↑ «WormBook: The online review of c. elegans biology»
- ↑ Hobert, O (2005), «Specification of the nervous system», in: The C. elegans Research Community, Wormbook, doi:10.1895/wormbook.1.12.1
- ↑ White, JG; Southgate E, Thomson JN, Brenner S (1986). «The Structure of the Nervous System of the Nematode Caenorhabditis elegans». Phil Trans Roy Soc London (Biol). 314: 1–340. doi:10.1098/rstb.1986.0056
- ↑ Parent, A; Carpenter MB (1995). Carpenter's Human Neuroanatomy. [S.l.]: Williams & Wilkins. pp. http://books.google.com/books?id=IJ5pAAAAMAAJ. ISBN 9780683067521
- ↑ Shu, DG; Conway Morris, S, Han, J., Zhang, Z-F., Yasui, K., Janvier, P., Chen, L., Zhang, X.-L., Liu, J.-N., Li, Y. and Liu, H.-Q. (2003). «Head and backbone of the Early Cambrian vertebrate Haikouichthys». Nature. 421: 526–529
- ↑ Striedter, GF (2005). Principles of Brain Evolution. [S.l.]: Sinauer Associates. pp. http://books.google.com/books?id=EPrJHQAACAAJ. ISBN 9780878938209
- ↑ Northcutt, RG (2008). «Forebrain evolution in bony fishes.». Brain Res Bull. 75: 191–205. PMID 18331871. doi:10.1016/j.brainresbull.2007.10.058
- ↑ Reiner, A; Yamamoto K, Karten HJ (2005). «Organization and evolution of the avian forebrain». Anat Rec a Discov Mol Cell Evol Biol. 287: 1080–102. PMID 16206213. Consultado em 12 de outubro de 2008[ligação inativa]
- ↑ Swaab, DF; Aminoff MJ, Boller F (2003). The Human Hypothalamus. [S.l.]: Elsevier. pp. http://books.google.com/books?id=Js81Pr1PmaAC. ISBN 9780444513571
- ↑ Jones, EG (1985). The Thalamus. [S.l.]: Plenum Press. pp. http://books.google.com/books?id=WMxqAAAAMAAJ. ISBN 9780306418563
- ↑ Saitoh, K; Ménard A, Grillner S (2007). «Tectal control of locomotion, steering, and eye movements in lamprey.». J Neurophysiol. 97: 3093–108. PMID 17303814. doi:10.1152/jn.00639.2006
- ↑ Puelles, L (2001). «Thoughts on the development, structure and evolution of the mammalian and avian telencephalic pallium.». Philos Trans R Soc Lond B Biol Sci. 356: 1583–98. PMID 11604125. doi:10.1098/rstb.2001.0973
- ↑ Salas, C; Broglio C, Rodríguez F (2003). «Evolution of forebrain and spatial cognition in vertebrates: conservation across diversity». Brain Behav Evol. 62: 72–82. doi:10.1159/000072438
- ↑ Grillner, S; Wallén P (2002). «Cellular bases of a vertebrate locomotor system-steering, intersegmental and segmental co-ordination and sensory control.». Brain Res Brain Res Rev. 40: 92–106. PMID 12589909. doi:10.1016/S0165-0173(02)00193-5
- ↑ Northcutt, RG (1981). «Evolution of the telencephalon in nonmammals.». Ann Rev Neurosci. 4: 301–50. PMID 7013637. doi:10.1146/annurev.ne.04.030181.001505
- ↑ Barton, RA; Harvey PH (2000). «Mosaic evolution of brain structure in mammals.». Nature. 405: 1055–8. PMID 10890446. doi:10.1038/35016580
- ↑ Pig brains kept alive outside bodies for first time could have major implications for humans Experiment raises concerns about ethics of future research into brains in laboratories por Josh Gabbatiss (2018)
- ↑ Calvin WH (1996). How Brains Think. [S.l.]: Basic Books. pp. http://books.google.com/books?id=z1r03ECL5A8C. ISBN 9780465072781
- ↑ Carew, Behavioral Neurobiology, Ch. 1
- ↑ Universidade de Rochester, ed. (7 de abril de 2016). «Study Reveals Brain's Finely Tuned System of Energy Supply». Consultado em 10 de dezembro de 2021
- ↑ Cooper et al, Biochemical Basis of Neuropharmacology
- ↑ Veja o artigo músculo
- ↑ Antle & Silver, 2005
Ligações externas
[editar | editar código-fonte]- «Cérebro X Caminhada - Pesquisas»
- «Brain Museum, Universidade de Wisconsin» (em inglês)
- «BrainWeb: Simulated Brain Database» (em inglês)
- «McConnell Brain Imaging Centre» (em inglês)